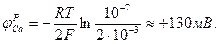Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизм генерации потенциала действия кардиомиоцита ⇐ ПредыдущаяСтр 9 из 9
Потенциал действия мышечной клетки сердца отличается от потенциала действия нервного волокна и клетки скелетной мышцы прежде всего длительностью возбуждения - деполяризации (рис).
Рис.. Потенциал действия кардиомиоцита Если длительность ПД аксона составляет 1 мс, клетки скелетной мышцы 2 - 3 мс, то длительность потенциала действия клетки сократительного миокрада желудочка и сердца составляет 250 - 300 мс. Это позволяет осуществить синхронное возбуждение и сокращение структур сердца для обеспечения выброса крови. Такие особенности ПД кардиомиоцита обеспечиваются распределением ионов внутри и снаружи клетки (рис.).
Рис.. Распределение концентрации ионов внутри и снаружи кардиомиоцита позвоночных (ммоль/л). Показаны К+- Na+- и Са2+- насосы, поддерживающие концентрации ионов на указанных уровнях; горизонтальными стрелками указаны направления пассивных потоков ионов при открытом состоянии соответствующих каналов, вертикальными - направление активного переноса ионов Распределение ионов К+ и Na+ в кардиомиоцитах близко к распределению этих ионов в скелетной мышце. Однако в кардиомиоците при формировании ПД и в процессе сокращения существенную роль играют и ионы Са2+. Их концентрация снаружи клетки составляет около 2 ммоль/л, но внутри клетки концентрация свободных ионов Са2+ очень мала: 10-4 ммоль/л. При сокращении концентрация свободных ионов Са2+ внутри клетки может возрастать до 10-8 ммоль/л, но в фазе реполяризации избыток этих ионов удаляется из клетки. Ионные насосы миокардиальных клеток. Сохранение ионного балланса в кардиомиоцитах обеспечивают К+- Na+- и Са2+-насосы, активно перекачивающие ионы Na+ и Са2+ наружу, а ионы К+ - внутрь клетки. Работу этих насосов обеспечивают ферменты К+- Na+- АТФаза и Са2+-АТФаза, находящиеся в сарколемме миокардиальных клеток. Плотность молекул К+- Na+-нacoca в мембране, оцениваемая по специфическому связыванию [3Н] - оуабаина, составляет около 1000 на 1 мкм2, то есть 1011 насосов на см2. Число циклов насоса оценивается ≈ 20 в секунду. Тогда на 1 см2 за одну секунду происходят 2 • 1012 циклов насосов. Так как за каждый цикл насос переносит 3 иона Na+, то всего переносится 6 • 1012 ионов за 1 с на 1 см2. Разделив этот результат на число Авогадро (6,02 • 1023 моль-1), получаем 10 • 10 12 моль/см2 • с, то есть по расчету через 1 см2 за 1 с насос перекачивает 10 пмоль ионов Na.
В покое проницаемость мембраны для ионов Na+ и Са2+ весьма мала: PNa/ Рк = 0,05; отношение РСа/ Рк также мало, мала и концентрация ионов Са2+ вне клетки. Поэтому потенциал покоя, как и в нервных волокнах, определяется в основном разностью концентраций ионов К+ по обе стороны клеточной мембраны. Потенциал действия клетки миокарда имеет три характерные фазы: деполяризация (I), плато (II) и реполяризация (III). I фаза — деполяризация, как и в аксоне, определяется резким ростом проницаемости мембраны для ионов натрия: Рк :PNa = 1: 20 в момент превышения φм порогового значения при возбуждении. Порог активации натриевых каналов примерно -60 мВ, а время жизни 1 - 2 мс и может доходить до 6 мс. II фаза - плато - характерна медленным спадом φм от пикового значения (= + 30 мВ) до нуля. В этой фазе одновременно работают два типа каналов - медленные кальциевые каналы и калиевые каналы. Кальциевые каналы имеют порог активации около -30 мВ, а время их жизни примерно 200 мс. В результате открывания кальциевых каналов возникает деполяризующий медленный входящий в клетку кальциевый ток: ICa=gCa(φM – φCa), где gCa - проводимость мембраны для ионов Са2+. Этот ток обеспечивается пассивным переносом в соответствии с градиентом электрохимического потенциала для ионов Са2+ (рис.). Равновесный кальциевый потенциал по уравнению Нернста:
Одновременно с ростом кальциевого тока растет проводимость для ионов калия gK, что приводит к возникновению вытекающего калиевого тока, ре поляризующего мембрану. Во II фазе gca уменьшается, a gK увеличивается (см. рис. 4.9), происходит постепенное выравнивание текущих навстречу друг другу токов, а потенциал мембраны φм понижается почти до нуля. Для II фазы характерно, что суммарный ток мембраны I стремится к 0.
Рис.. Изменение проводимостей для ионов Na+, Ca2+, К+ при возбуждении каридомиоцита III фаза — реполяризация - характеризуется закрытием кальциевых каналов, ростом величины gK и усилением выходящего тока К+. Для кальциевого канала, так же как и для натриевого, предполагается существование активирующих и инактивирующих частиц, состояние которых описывается некоторыми параметрами d и f соответственно. Тогда проводимость канала gCa в уравнении:
gCa= gCa∙d∙f, где gCa — максимальная проводимость открытого кальциевого канала. Процессы возбуждения кардиомиоцита изучаются с помощью ряда специальных методов.Один из них - это метод блокаторов (антагонистов) ионов кальция. Были найдены специфические блокаторы кальциевого тока в миоците: препараты Д-600, верапамил, катионы металлов La3+, Mn2+ и некоторые другие. Эти вещества прекращают доступ кальция внутрь клетки и тем самым изменяют величину, и форму потенциала действия. Интересно отметить, что кальциевые каналы не блокируются тетродотоксином (блокатором ионов Na+), что дает основание допускать существование в кардиомиоцитах отдельных кальциевых каналов. Второй метод - люминесцентный анализ. Он позволяет регистрировать в эксперименте перенос ионов кальция с помощью, белка экворина, получаемого из светящихся медуз. Особенность этого белка заключается в том, что, обладая высоким сродством к ионам Са2+, он люминесцирует в их присутствии. Экворин S вводится в препарат сердечной мышцы, и с помощью специальной оптической аппаратуры регистрируется изменение интенсивности свечения во времени. Полученные результаты пoзволяют описать процессы переноса ионов кальция при генерации потенциала действия в мышце сердца. Распределение ионов кальция по сердечной мышце в норме и патологии изучается с помощью метода радионуклидной диагностики. Для этого используют радиоактивный изотоп кальция – Ca2+, β - излучение которого регистрируется сканерами. БИОФИЗИКА КЛЕТОК И ОРГАНОВ
|
||||||
|
|
Последнее изменение этой страницы: 2017-02-07; просмотров: 450; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.24.134 (0.008 с.) |