Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Принципы флористического и фаунистического районирования суши. Понятие «эндемизм»Содержание книги
Поиск на нашем сайте
Биота суши в широком смысле делится на сухопутную и биоту континентальных водоемов. Наиболее резко обособленность биоты континентальных водоемов выражена у организмов, весь жизненный цикл которых проходит в водной среде (моллюсков, ракообразных, рыб и др.). У постоянноводных организмов континентальных водоемов историко-провинциальное распределение по бассейнам выражена резче, чем связь с геопоясными, зональными и секторными подразделениями суши. Локальный эндемизм, иногда очень резкий, присущ глубоким долгоживущим озерам. Для суши наиболее разработаны флористические и фаунистические районирования. А также ботаникогеографическое. Флористическое и фаунистическое (зоогеографическое) районирования опираются исключительно на анализ ареалов, регионы выделяются и их ранг определяется по обилию и рангу эндемиков и по соотношению видов с разными типами ареалов. При объединении выделов низкого ранга в более высокий эндемизм складывается из эндемиков составляющих единиц и эндемиков более высокой единицы, охватывающей большинство составляющих низших выделов. При учете выделов высоких рангов в число их эндемиков попадают и виды не характеризующиеся как эндемики каких-либо выделов более низкого ранга. В ботанике отдельно существуют флористические и геоботанические районирования часто рассматривающиеся как параллельные и необъединимые, но создаются также и схемы, объединяющие оба направления. В зоогеографии суши преобладает фаунистическое направление. Часть несовпадений флористических, фаунистических и геоботанических делений связана с неполным учетом фауны (только позвоночные) с крайностями принципов флористического и фаунистического районирования, ориентирующегося преимущественно на эндемиков, с невзвешенным учетом молодых и реликтовых эндемиков и др. Один из первых опытов создания флористической системы суши принадлежит датскому ботанику Д. Скоу, который в 1823 г. выделил 25 царств, часть из которых он подразделил на провинции. В качестве критерия при построении флористической системы Д. Скоу учитывал, в частности, степень эндемизма таксонов разного ранга, присущую тем или иным территориям. Этот критерий сохраняет свое значение и поныне. Большой вклад в развитие флористического районирования внесли А.Энглер (1924), Л.Дильс (1918), Р.Гуд (1946), А.И.Толмачев (1974) и др. В дальнейшем изложении будем придерживаться системы районирования, разработанной А. Л. Тахтаджяном (1978). Высшая единица флористического районирования - царство. Ранг царства присваивается регионам, характеризующимся максимальным своеобразием флоры в целом и наличием эндемичных таксонов высокого ранга (семейств, подсемейств) в сочетании с очень высоким числом эндемиков родового и видового ранга. Всего выделяют шесть царств (табл. 3). Царства подразделяются на полцарства и области, которые характеризуются высоким родовым и видовым эндемизмом; кроме того, каждой области присущ определенный набор семейств, занимающих в ней лидирующее положение. Всего выделяют 35 областей, которые подразделяются на провинции. Низшей хорологической единицей является округ. Таблица 4 Фаунистические регионы суши
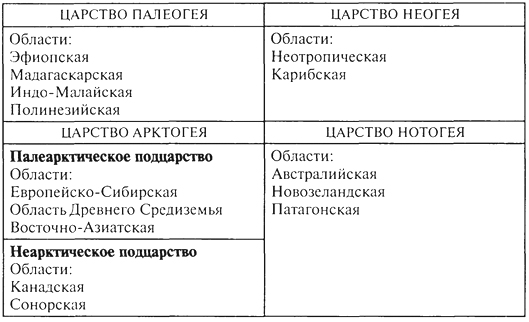
Принятая в учебнике система фаунистических регионов суши представляет собой модернизированную схему П.Склэтера - А.Уоллеса (табл. 4, рис. 55).
Одной из важнейших особенностей фауны является эндемизм. Эндемичными систематическими единицами называются виды, роды и другие таксономические единицы, которые встречаются только в какой-то определенной области и нигде более. В силу этого эндемики представляют специфический компонент любой фауны и абсолютно отличают ее от других фаун. Если более широко распространенные виды указывают на связи между фаунами, то эндемики резко отличают их друг от друга. Количество эндемиков в различных фаунах неодинаково. Наиболее высок процент эндемизма в островных фаунах, а на континентах — в районах с сильно расчлененным рельефом, т. е. в горных странах, поскольку географическая изоляция — необходимое условие процесса видообразования. Для образования эндемичных видов требуется не только географическая изоляция, но и продолжительное время. Длительное и непрерывное развитие фауны благоприятствует возникновению новых видов, в то время как быстрые изменения условий существования, наоборот, ведут к вымиранию одних и расселению других. Обилие эндемиков отражает длительность развития фаун. Различия между эндемичными и неэндемичными компонентами фауны не абсолютны. В большинстве фаун есть так называемые субэндемики, или почти эндемичные таксоны (роды, виды).
Флористическое районирование. Общая схема.
Флористические цартсва выделяются неповторимым, своеобразным обликом. Совершенно обязательно есть определенное число эндемичных семейств
Эпифиты – растения использующие в качестве субстрата ствол другого растения. Регионы различны Царства делятся на области. Каждая область характеризуется определенным обликом растительности. Всего 6 флористических царств. Голарктика Неотропическое и Палеотропическое Капское царство Гоантарктическое Автралийское
Фильтры и мосты. Островные флоры.
Голарктика 3 Подцарства. Бореальное Хвойные, 4 многочисленных вида. Ель, сосна,лиственница, пихта. В американской части голарктики добавляются тцуга и туя. Древнее седиземноморское подцарство Мадрианское подцарство.
Фаунистическое районирование суши Высшая единица фаунистического районирования – царство. Ранг царства присваивается регионам, характеризующимся максимальным своеобразием фауны в целом и наличием эндемичных таксонов высокого ранга (семейств) в сочетании с очень высоким числом эндемиков родового и видового ранга.
Фаунистические царства: Нотогея (Австралийское+ Антарктическая область) Палеогея (Индо-Африканское) Неогея (Южная Америка) о ней писал Симпсон «Великолепная изоляция» Арктогея 37. Планетарный, региональный и топологический (ландшафтный) уровни дифференциации живого покрова суши. Зональные и интразональные типы сообществ.
Одним из основных понятий в физической географии является природная зона. Природные зоны — это крупные подразделения географической оболочки Земли, закономерно и в определенном порядке сменяющие друг друга в зависимости от климатических факторов, главным образом от соотношения тепла и влаги. Смена зон происходит в меридиональном направлении, от полюсов к экватору. В каждой зоне составляющие ее природные компоненты, прежде всего климат, почвы, сообщества растений и животных (биоценозы), имеют типичные особенности. Для сообществ климатические и почвенные условия в пределах зоны являются зональными экологическими факторами. Названия зонам обычно даются по растительности, поскольку растительность, как отмечалось, служит ведущим компонентом биоценоза. Сообщества бывают зональные и азональные (незональные). Зональные занимают в пределах зоны плакорные местообитания - - выровненные, хорошо дренируемые водораздельные пространства с супесчаными и суглинистыми почвами без болотообразовательных процессов. Только в этих местообитаниях в полной мере проявляется зависимость биоценозов, в первую очередь фитоценозов, от климата данной збны. Сообщества за пределами плакоров зоны получили название азональных. Среди них выделяются интразональные и экстразональные. Под интразональными понимаются сообщества, распространенные в одной или нескольких зонах на отдельных участках. Их существование определяется в основном почвенно-грунтовыми условиями, отличающимися от плакорных. Примерами интразональных сообществ могут служить сообщества верховых болот и сосняков на песчаных почвах в лесной зоне, солончаков и солонцов в степной и пустынной зонах, луговые сообщества пойм. Нередко интразональные сообщества преобладают над зональными, например болотные массивы в Западной Сибири, однако интразональные сообщества всегда формируются под ведущим влиянием зональных географических условий. 38. Первичные и вторичные сукцессии. Их стадии. Антропогенные сукцессии в тайге.
Происходящие во времени смены одного биоценоза другим получили название сукцессии. Одно из важнейших свойств биоценоза - способность реагировать на направленное изменение условий существования последовательными сменами. Так, под воздействием непрерывного, медленного, но неуклонного повышения уровня грунтовых вод на какой-то территории происходят изменения влажности почв, миграции солей, условий аэрации, что существенным образом сказывается на корнеобитаемом слое. В связи с этим происходит изменение ценотически сложившихся отношений в биоценозе, численность одних видов снижается вплоть до их исчезновения, происходит постепенное внедрение новых видов извне, с течением времени возникает новый биоценоз. Направленная и непрерывная последовательность появления и исчезновения популяций разных видов, изменение видового состава биоценоза во времени, приводящая к последовательной смене одного биоценоза другим, и есть сукцессия, а отдельные биоценозы, соответствующие разным этапам развития биоценозов, - это стадии сукцессии. Глубокая разработка теории сукцессии связана с именами американских ученых - Г. Коулса и Ф. Клементса; большой вклад в развитие этой теории внесли крупнейшие отечественные ученые - Г.Ф. Морозов, И. К. Пачоский, Г. Н. Высоцкий и особенно В. Н. Сукачев. Наиболее полно изучены сукцессии растительности, что объясняется огромной ролью растений в формировании структуры биоценоза и развитии сукцессии, их способностью в известной степени направлять ее ход, контролируя возможности существования для других организмов. Это не исключает роли животных, особенно фитофагов и землероев, в развитии тех или иных стадий сукцессии, их воздействия на численность и существование тех или иных видов. Сукцессии разномасштабны как по площади, которую они охватывают, так и по продолжительности. Это смены, например, при зарастании временных водоемов, склонов молодых оврагов, гарей, залежей, сукцессии на выбросах земли из нор кротов, сусликов. Длительные вековые смены связаны с крупномасштабными климатическими изменениями. Принципиально важно разделение сукцессии на первичные и вторичные. Первичные сукцессииразвиваются на субстратах, где ранее жизнь отсутствовала, это сопряженные процессы почвообразования и постепенного формирования растительности и животного населения, очень медленные, протекающие столетиями. Таковы смены растительности на породах, обнажившихся после таяния ледников или на побережьях вследствие отступания моря. Так, на обнажившихся скальных породах развитие биоценоза начинается с поселения накипных лишайников или пионерных видов в трещинах скал. Это, как правило, очень длительная стадия, для которой характерно также присутствие водорослей, некоторых простейших, насекомых и клещей. По мере разрушения твердой породы и постепенного образования тонкого рыхлого слоя поселяются другие группы лишайников, различные виды мхов, увеличивается разнообразие животных, продолжается формирование почвы, с развитием которой происходит вселение сосудистых растений, идет обогащение животного населения. Таким образом, в ходе первичной сукцессии идет процесс первоначального становления биоценоза, связанный с заселением нового пространства пионерными формами жизни, отбором видов в процессе освоения конкретных условий среды и конкуренцией за средства жизни - эцезис. Важная движущая сила сукцессии в данном случае - изменение почвы первыми поселенцами. И куропаточья трава, и ольха имеют симбионтов, фиксирующих атмосферный азот, что приводит к накоплению в почве его запасов. Ольха также способствует сильному подкислению почвы, снижая за 50 лет рН в ее верхних горизонтах приблизительно с 8,0 до 5,0. После этого ситхинская ель, используя накопленный азот, может поселяться и замещать ольху. Постепенное накопление в почве углерода ведет к развитию мелкокомковатой структуры, повышению аэрации и влагоëмкости субстрата. На хуже дренированных склонах сукцессия не всегда заканчивается елово-тсуговым лесом. Во влажных местах, с худшим дренажем, бурно развиваются сфагновые мхи, накапливающие воду и сильно подкисляющие почву, которая испытывает дефицит кислорода; при этом большинство деревьев гибнет. Единственный вид, способный выдержать плохую аэрацию субстрата возникающего болота, - сосна скрученная. Таким образом, в развитии сукцессии важное значение приобретают экологические различия; формирование растительности на поздних стадиях сукцессии контролируется почвенно-геоморфологическими условиями. Вторичные сукцессии - это смены биоценозов, начинающиеся от сформированной растительности. Там, где есть почвы с запасом семян, спор, различных вегетативных зачатков, развиваются вторичные сукцессии, как правило, протекающие намного быстрее первичных. Вторичные сукцессии развиваются под воздействием внешних по отношению к биоценозу факторов (изменение уровня грунтовых вод, подтопление, накопление токсичных солей в верхних почвенных горизонтах, выпас, вырубки, пожары) и в результате деятельности биоты. Если внешнее воздействие вызывает полное или почти полное уничтожение растительности, то вторичная сукцессия развивается в направлении восстановления нарушенного (уничтоженного) биоценоза. Так, на месте пожара или вырубки начинается восстановительная сукцессия, смена биоценозов идет в направлении к состоянию, близкому к исходному, существовавшему до нарушения. Этот процесс восстановления в ходе сукцессии исходных биоценозов получил название демутации. В ряде случаев, например под воздействием выпаса, происходит постепенная смена биоценозов, сопровождающаяся упрощением структуры, обеднением видового состава и на крайних стадиях характеризующаяся почти полным исчезновением растительности (сильно выбитые пастбища). Этот процесс известен под названиемдигрессия.
|
||||
|
|
Последнее изменение этой страницы: 2017-01-25; просмотров: 862; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.189.143.1 (0.008 с.) |