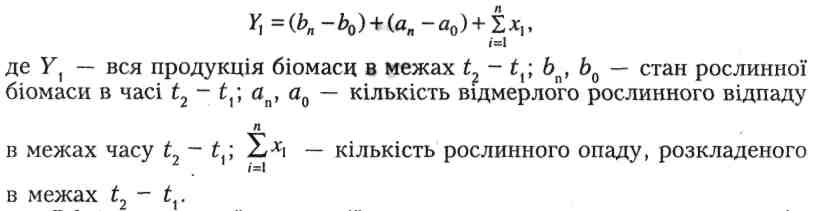Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Потік енергії та продуктивність екосистемиСодержание книги Поиск на нашем сайте
Крупні сучасні екосистеми настільки складні, що їх сутність, можливо, найкраще вивчати враховуючи стадії еволюції життя на нашій планеті, на якій виникає дана екосистема. Відомо, що багато органічних сполук синтезовано в природі без втручання живих організмів, чиї форми життя народжувалися в органічному "бульйоні" дрібних водоймищ. Ці водоймища часто висихали, а з ними зневоджувалися і тіла живих організмів. Завдяки високій життєздатності вони могли тривалий час очікувати наступного обводнення свого колишнього місцезростання або ж "перебиратися" за допомогою вітру в інше водоймище з подібним органічним "бульйоном". Потрапивши в сприятливі умови, ця життєва форма ставала знову активною (Рис.1.).
Рис.1. Перша "екосистема". Розглядаючи таке первісне водоймище, Дж. Казенс моделює екосистему. Він припускає, що основний корм (джерело енергії) якоїсь форми життя – органічна сполука А, що розпалася лише до рівня сполуки В (яка через це має менше енергії). Тепер уявімо іншу життєву форму, котра використовує як корм органічну сполуку В, розщеплюючи її на неорганічні речовини, і ми, – заключає вчений, – одержимо основи екосистеми, яка включає: 1. Повне перетворення органічної речовини А в неорганічну речовину В з виділенням енергії. 2. Один функціональний зв'язок між формами життя. Друга істота залежить від першої, яка виробляє сполуку В. Змодельована первісна екосистема залежить від акумулюючої речовини А і достатньо ефективного механізму розсіювання, щоб забезпечити появу обох видів у місцях такої акумуляції з регулярними інтервалами. У процесі еволюції фотосинтезуючих організмів, можливо, мільйони років опісля, потенціал біосинтезу органічної речовини виріс на багато порядків, а абіотичний синтез органічних сполук став несуттєвим як джерело енергії. Отже, перші екосистеми, як зазначено вище, були гетеротрофними, тобто залежали від забезпечення органічною речовиною, водночас як абсолютна більшість сучасних екосистем – автотрофні і їх існування залежить від здатності зелених рослин поглинати сонячну енергію. Незважаючи на те, – підкреслює Дж.Казенс, – що еволюція видів рослин і тварин привела до збільшення розмірів і структурної складності як індивідуумів, так і біогеоценозів, фундаментальні процеси залишилися ті ж, а саме: 1. Синтез органічної речовини, переважно біосинтез матеріалу нових клітин (біомаси); енергія, яку одержує екосистема, – це головним чином сонячне випромінювання. 2. Біологічне розщеплення біомаси і мертва органічна речовина (МОР), одержувані з втратою енергії у вигляді тепла і виділенням неорганічних хімічних речовин. Оскільки енергія не втрачається, а лише трансформується, бюджет енергії має бути збалансованим. Використовуючи термін енергетика, – зауважує Дж.Казенс, – енергію розглядають як таку, що: а) вливається в екосистему, б) переливається між сховищами органічних речовин, в) виливається з неї у вигляді тепла. Однак потік енергії, який проходить через систему, є лише побічним продуктом двох головних процесів, про які згадано вище. Біологічне розщеплення – це вивільнення неорганічних хімічних сполук, необхідних для наступного біосинтезу автотрофними організмами. Біологічний розклад зумовлює циркуляцію поживних речовин, а саме елементів всередині системи. Якщо настає рівновага між втратою і накопиченням поживних речовин в екосистемі, то система потенційно здатна функціонувати, доки триває надходження енергії [1].
Потік енергії в популяції Р.Дажо наводить приклад перенесення енергії в популяції антилопи (Уганда), чисельність якої оцінювалась в 15000 особин масою 2174 кг/км кв. Популяція займала 400 км кв площі місцевого заповідника. За результатами досліджень вчені одержали такі дані (ккал/м кв/рік): З'їдений корм – 74,1 Потік енергії – 62,4 Метаболізм – 61,6 Приріст популяції – 0,81 Таким чином, на приріст популяції витрачено мізерну кількість енергії – усього близько 1%. Приріст відповідає утворенню 577 кг живої речовини на 1 км кв. Р.Дажо для порівняння наводить приклад проходження енергії через популяції чотирьох травоїдних видів: полівки, слона, віргінського оленя і угандійської антилопи (ккал/м кв/рік): Полівка – 17,5 Слон – 23,3 Олень – 43,1 Антилопа – 62,4 Із вивчених ссавців популяція угандійської антилопи вирізняється тим, що через неї проходить найбільший потік енергії. Це пов'язане з великими розмірами тварини і щільністю популяції. Антилопа використовує 10% первинної продукції, тоді як полівка – 2%, олень – 4,5%. Найвищий потік енергії серед тварин усього світу зафіксовано у цикадки Prokelesia (Homoptera) із прибережних боліт Джорджії (США) – 275 ккал/м кв/рік. Цікаві дані наводить Ю.Одум (1962), який зі співробітниками вивчав потік енергії, що проходить через популяції травоїдних прямокрилих – кобилок і цвіркуна (штат Джорджія). Виявилось, що маса листя, яке з'їли прямокрилі, становить 85% чистої продуктивності поля, тоді як частка зерна, з'їдженого хребетними, не перевищує 7%. Крім того, відсоток споживання корму більшою мірою залежить від харчового раціону.
Використання енергії зерноїдними і травоїдними тваринами
Як бачимо, відсоток використання корму травоїдними прямокрилими (від 2 до 7%) є досить низьким. Зерноїдні хребетні відзначаються значно вищим відсотком використання енергії (10-50%). Наведені дані підтверджують теорію Хейрстона (1960), згідно з якою величина популяції травоїдних тварин, як правило, не пов'язана з відсутністю корму, тоді як у хижаків, детритоїдних і зерноїдних тварин вона часто визначається саме цим. Взимку, коли корму стає мало, вона може стати для зерноїдних, які посідають в екосистемі проміжне місце між м'ясоїдними і травоїдними, одним з лімітуючих факторів. Із енергетикою обміну речовин пов'язані плодовитість, тривалість життя, розмір тіла, раціон і рух популяцій (рис. 2.). Для підтримки свого існування крупним тваринам потрібна більша кількість речовин та енергії, ніж дрібнішим, а щоб одержати цю кількість, вони вимушені переміщатися ширшими просторами, ніж тварини невеликих розмірів, які споживають подібний корм. Оскільки корм травоїдних тварин є звичайно в надлишку, вони не займають великих територій. Хижаки ж, а також ті травоїдні, які вимушені шукати свій корм (наприклад, насіння або плоди), витрачають багато енергії. Перша група – то "женці", а друга – "мисливці". "Мисливці" за своєю природою "територіальники", тобто захищають свою територію, на що витрачають чимало енергії, "женці" майже не охороняють "своє поле".
Рис.2. Схема. взаємозв'язків факторів, які впливають на продукційні процеси (кругообіг енергії і матерії) популяції.
Метаболічна вартість руху залежить як від його способу, так і від розміру тіла тварини. Зусилля на переміщення одиниці тіла на деяку стандартну віддаль для крупних тварин в дійсності менше, ніж для дрібних. Пересування по суші вимагає найбільших затрат, політ – середніх, а найекономнішим є плавання [1]. 2.2. Потік енергії через популяцію Особина впродовж активного життя непомітно пов'язана з середовищем якраз потоком енергії і матерії, котрий надходить разом з кормом із середовища до особини, в якій ці два компоненти і матерія нагромаджуються у вигляді біомаси. Одночасно частина енергії та матерії після певних перетворень особиною надходить до середовища у вигляді продуктів обміну речовин і тепла. Цей процес має назву біологічної продуктивності організму. Той самий процес, що й у випадку однієї особини, вирішує долю життя, приросту біомаси, народжуваності і тривалості життя конкретної популяції рослин чи тварин. Потік енергії і матерії через популяцію називають біологічною продукцією популяції. Для того, щоб створити нову матерію – продукцію, необхідний корм (С). Наступний процес засвоєння особиною частини матерії у вигляді корму становить брутто продукції, або ж асиміляції (А). Частина засвоєної продукції йде на побудову тіла особини, яке називають біомасою (продукція нетто Р). Наявна в кормі енергія використовується на життєві процеси особини (пересування, втеча, погоня, пережовування корму і т.п.), а також розсіюється у вигляді теплової, хімічної (наприклад, газовиділення), світлової, електричної енергії. Оскільки всі організми беруть енергію з окисленої органічної субстанції (вуглецю), її мірилом є інтенсивність дихання особин (респірація (R), від лат. "респіро" – дихання). У тварин неперетравлений корм, а також фекальні рештки (кал, сеча або урина), які виділяються в середовище (FU), є не що інше, як різниця між спожитою їжею і асиміляцією (FU=(C-A)). Подібно до тварин рослини виділяють корінням і надземними органами продукти асиміляції: вуглекислий газ, токсичні метаболіти, а також невикористані метаболізмом сполуки. Як ми вже знаємо, ці виділення можуть бути для інших організмів шкідливими (алелопатія), корисними (біостимуляція) або ж можуть не приносити ні шкоди, ні користі. Крім згаданих виділень, які утворюються внаслідок засвоєння корму, тварини і рослини виділяють ще різноманітні слизи. Всі вони разом (фекалії, сеча, слиз і т.п.) і будуть становити непродуктивні відходи життєдіяльності особин (FU). Розглянемо загальні рівняння енергетичного балансу для особин і популяції, сформульовані польським біологом К.Петрусевичем (1978): С = Р + R + FU; А = Р + R = С - FU; P = A - R = C - FU - R. Однак слід зауважити, що ці рівняння енергетичного балансу певною мірою спрощені і мають теоретичний характер. Якщо б навіть вдалося в лабораторних умовах контролювати всі названі параметри, навряд чи можна було б домогтися такого ідеального балансу [1]. 2.3. Продуктивність популяції Розглядаючи ідею продуктивності в екології, К. Петрусевич наводить такий приклад. Якщо в певний момент Т1, маємо, наприклад, 10 особин (N1), а в момент Т2 їх виявилося N2=20 особин, то це означає, що різниця N2 - N1 = ΔN = 10. Це може бути у випадку народження 10 особин і відсутності відпаду або у випадку народження 25 особин і загибелі 15, а також при народженні 100 особин і загибелі 90. Таким чином, різниця стану, підкреслює К. Петрусевич, говорить нам дуже мало або й зовсім нічого про те, який був доплив особин до популяції. Отже, збільшення особин у популяції і є її продукцією, вираженою в особинах. Однак кожна особина має свою масу, а отже, сумарна маса особин популяції являє собою масу популяції, яка може виражатися в одиницях маси – тоннах, центнерах, кілограмах, грамах чи міліграмах. У першому і другому випадках у енергетичному балансі особин продукція складається з двох елементів: приросту чи зменшення біомаси і одночасно продукції потомства. Причини спаду динаміки є різні: натуральна смертність, втрати, які несе популяція внаслідок її експлуатації іншими популяціями на певному трофічному щаблі. Часто доплив і відплив з популяції зображають у вигляді водяного стовпа з різними варіантами вхідного і вихідного отворів (рис. 3.). Надходження води до резервуару (р) ілюструє продукцію популяції, а розхід (е) – елімінацію (втрати, відпад, винесення особин за межі популяції). Висота водяного стовпа (h) ілюструє щільність популяції. В такому випадку втрати e є функцією продукції і щільності популяції: е = f (p,h). Коли потік є сталий, то ширина вхідного і вихідного отворів може бути чи великою, чи малою, а пропорція водяних стовпів залишається однаковою h1 = h2. Цей випадок добре ілюструють наведені на початку розділу варіанти зміни чисельності популяції при загальному її збільшенні на 10 особин. В іншому випадку врівноважений стан забезпечується більшим допливом і меншим відпливом біомаси. Як бачимо, цей багатокомпонентний і динамічний процес тяжко вкласти в рамки простого рівняння, яке краще всього могло б розв'язуватися в умовах, коли можна було б припинити відпад і провести обміри біомаси. Однак у природі (як і в лабораторії) цей динамічний процес зупинити не можна, як не можна зупинити життя.
Рис.3. Врівноваження взаємного стану біомаси (В), продукції (р) і відпаду (е). Цей стан біомаси може утримуватися як при високому рівні продукції системи, так і при низькому. У звичайних умовах загальну продукцію популяції можна зобразити приростом біомаси (ΔВ) і елімінацією або відпадом (Е):
Зауважимо, що найвищий рівень продуктивності популяції у випадку найменшого відпаду. В природі трапляються такі випадки в агроценозах, коли кількість організмів експлуатуючої популяції усувається методами хімічного захисту, забезпечуючи можливість максимального приросту експлуатованої популяції. В рівнянні Р.Вігерта, Ф.Еванса (1964) оцінюється первинна продукція з урахуванням впливу, який на неї має явище детрифікації рослинного матеріалу:
Обмір первинної продукції природних угруповань охоплює, крім оцінки станів біомаси, кількість усього матеріалу та з'ясування темпів розкладу мертвого матеріалу. В популяціях, які в річному циклі дають лише одне покоління, а межі розроджуваності є короткі, оцінка продукції може бути проведена в межах початкового виходу особин (N0) та їх наступної редукції. Прикладом такого виду є бурякова стонка, яка проходить чотири личинкові стадії, під час яких живе на листях буряка, годуючись ними. Стадія лялечки перебуває в ґрунті, а доросла особина знову живе на листях буряка аж до настання холодів, після чого заривається в землю на зимування. Оцінка повної чисельності особин у популяції можлива з урахуванням відкладених яєць. Оцінка росту чисельності у різних стадіях личинки можлива на підставі кривої редукції (рис.4). Продукція, створена в окремих стадіях личинки, є функцією термічних умов середовища. Загальна продукція популяції в стадії личинки (Р) дорівнює
де ΔВ – приріст біомаси на даній стадії личинки; N0, N1 – початкова і кінцева чисельність особин у даному стані личинки; L1, L4 – чергові стадії личинок. Загальна продукція, яка обіймає повний цикл розвитку виду, дає достатньо даних про приріст біомаси і стан енергії, закладеної в літньому жуку, який з'являється в серпні й інтенсивно жирує на посадках буряка до викопування або до повного з'їдання листя. У популяціях, в яких декілька поколінь виступає одночленно, а розвиток має тривалий характер, основним способом визначення продукції є дослідження обороту особинами.
Рис. 4. Хід природного розмноження бурякової стонки на посівах буряка в західній частині Польщі: 1 – в кінці червня, 2 – в липні, 3 – на початку серпня, L1-L4 – чергова стадія личинки. Оборот, як і інші параметри продуктивності (Р, A, R, С, FU), розглядають у певних часових межах (звичайно за рік, за сезон). Звідси витікає загальне визначення продукції: це повна кількість органічної матерії, створена в даному (певному) часі, не використана на распірацію (P=A-R), незалежно від того, чи залишилася ця матерія в популяції на кінець даного часу (Р = дельтаВ + Е), чи буде винесена. Наведемо п'ять різних варіантів математичного виразу продукції за К. Петрусевичем: 1) Р = А - R (сальдо нетто між допливом енергії і коштом на її утримання); 2) Р = С - R - FU (сальдо нетто вжитого корму і фекалій разом з уринами (сечею) та кошту утримання); 3) Р = ΔВ + Е (приріст біомаси разом з відпадом); 4) Р = Pr + Pg (продукція зростає внаслідок росту тіл і розроджуваності); 5) Р = B·Q (середня біомаса в обороті). Отже, біомаса, створена популяцією за одиницю часу, є продукція нетто цієї популяції. На цю продукцію накладаються два процеси: приріст чисельності особин, випродукованих популяцією, а також приріст біомаси цих особин [1]. 2.4. Експлуатація популяції Проблема експлуатації популяції має як теоретичне, так і практичне значення. Це, зокрема, створення основ охорони видів, яким загрожує винищення, а також ведення мисливського господарства і рибальства. Якщо йдеться про види, котрі потребують посиленої охорони, то виходять з положення, що скорочення чисельності, а також загроза повного винищення популяції є наслідком експлуатації виду консументами, які її використовують як корм. Часто такі проблеми вирішували обмеженням популяції-експлуататора, але ці дії не завжди приводили до позитивного наслідку. Наприклад, в одному з районів штату Арізона (США) у 1905 р. налічувалось близько 4 тис. оленів і дві популяції хижаків – пуми і вовка, які експлуатували популяцію оленя як корм. Однак інтенсивне полювання на цих хижаків призвело до того, що їхня чисельність, починаючи приблизно з 1918 p., стала скорочуватися. Буквально впродовж десятка років поголів'я оленів зросло до 100 тис, але потім почало стрімко падати і в 1939 р. налічувало лише 10 тис. особин. Однією з причин такого спаду стало порушення рівноваги в пасовищній екосистемі: витоптані травостої не могли витримати такий потужний прес та задовольнити кормові потреби дорослого поголів'я, кількість якого колись успішно регулювали хижаки. В експлуатаційних рядах популяцій спостерігається і такий: рогіз-карась-щука (хижак першого порядку) – людина (хижак другого порядку). Наслідки експлуатації одної популяції іншою можна представити як нетто кормового ланцюга:
Підставивши у формулу дані у калорійному обчисленні, одержимо продуктивність нетто кормового ланцюга. Компенсувати збитки від експлуатації популяція може трьома шляхами: а) підвищенням народжуваності; б) зниженням смертності; в) підвищенням біомаси в циклі розвитку особин [1].
|
||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-23; просмотров: 470; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.63.176 (0.013 с.) |