Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
А (дополнительная). Взаимоотношения хищника и жертвы в экологическом и эволюционном масштабах времени.Содержание книги
Поиск на нашем сайте
Одной из приоритетных задач современной биологии, несомненно, является создание обобщенной теории эволюции экосистем. При этом у экологов может возникнуть сильный соблазн начать ее строить в одиночку, опираясь исключительно на данные по динамике современных сообществ и игнорируя "измышления" палеонтологов. А почему бы и нет? Есть тому пример: эволюцию всех конкретных таксонов животных и растений реконструировали, как всем известно, при самом минимальном участии палеонтологов, опираясь почти исключительно на данные сравнительной анатомии. Для этого достаточно сходные рецентные организмы располагают в виде последовательностей, трактуемых как отражение реальной премственности в ряду "предок-потомок" (например, тупайя - лемур - низшая обезьяна - человекообразная обезьяна - человек), и уже потом эти филетические ряды дополняют ископаемыми членами (в нашем случае - австралопитеком, питекантропом и неандертальцем).
Хотя злые языки из числа антиэволюционистов и утверждают, будто филетические ряды мало чем отличаются от известной карикатуры XIX века, изображающей "Чудесное превращение короля Луи-Филиппа в грушу", это, разумеется, не так. Настоящая проблема в другом: при реконструкции исторических последовательностей косвенный, "сравнительно-анатомический", метод действительно предпочтителен (поскольку метод прямой, "палеонтологический", имеет крупный встроенный дефект в виде принципиальной неполноты летописи), однако он годится не для всякого типа биологических объектов. А именно - он приложим лишь к тем объектам, которые хотя бы в принципе обладают филогенезом, т.е. историческим развитием; иными словами - система должна обладать хорошей памятью относительно своих предшествующих исторических состояний.
В случае таксона память эта обеспечивается передаваемым от предка к потомку "текстом" в виде нуклеотидной последовательности. А вот экосистема подобного наследственного субстрата лишена; она обладает лишь элементами онтогенеза, т.е. индивидуального развития (экогенетические сукцессии, о которых речь пойдет в главе 12), но филогенеза у нее, судя по всему, не бывает в принципе. Так что для изучения истории экосистем "сравнительно-анатомический метод" не годится, и палеонтология, как это ни парадоксально, неизбежно будет играть здесь существенно большую роль, чем при реконстукции филогенезов отдельных таксонов.
Итак, создавать теорию эволюции экосистем должны, на паритетных началах, экологи и палеонтологи, причем первым, по идее, следует создавать основу для актуалистической интерпретации тех фактов, что накоплены вторыми. К сожалению, в процессе такого "заочного сотрудничества" между экологами и палеонтологами часто возникают недоразумения, связанные с неверным пониманием границ применимости каждого из используемых ими методов. Одну из таких "конфликтных ситуаций" можно разобрать на примере взаимоотношений хищника и жертвы в докембрийских планктонных экосистемах) [17].
В современной экологии существует достаточно хорошо обоснованная (в том числе и математическими моделями) концепция оптимального питания хищника. Согласно ей, консумент стремится выбирать пищевые объекты таким образом, чтобы разность между энергосодержанием пищи и суммарными энергозатратами на ее добывание оказывалась максимальной. Для каждого конкретного хищника существует определенный размерный оптимум жертв, с наибольшей частотой поедаемых им (лев питается в основном антилопами, хотя в составе его добычи могут оказаться и буйвол, и кролик). В свою очередь, у жертвы имеется целый ряд эволюционных стратегий, защищающих ее популяции от выедания (увеличение скорости размножения, приобретение криптической окраски или ядовитости, и т.д.); экспериментально показано, что одной из таких стратегий является уход за пределы размерного оптимума.
Из наблюдений над сезонными изменениями в популяциях планктонных ракообразных и коловраток было установлено, что это действительно так; при этом жертва по-разному защищается от тех хищников, что лишь немного превосходят ее по величине, и тех, что относятся к следующему размерному классу. Защитой от врагов первого типа (выедающих особей среднего и мелкого размера) является увеличение собственых размеров жертвы за пределы оптимума выедания. Против врагов второго типа (выедающих самых крупных из жертв) такая стратегия бесполезна, здесь надо уменьшать свои размеры. Защитой же от комбинированного воздействия хищников обоих типов может служить изменение формы тела - образование шипов и прозрачных выростов кутикулы (их периодическое появление в популяциях дафний - хорошо известный факт). Это позволяет увеличить реальные размеры (создав помехи для врагов первого типа), сохраняя при этом возможно меньшую заметность для врагов второго типа. Основываясь на этой экологической концепции, а также на эмпирических данных о распространении в геологической истории основных размерных классов фитопланктона (до 16 мкм, 16-60 мкм, 60-200 мкм, свыше 200 мкм) и его морфологических групп (оболочка гладкая или с выростами), М.Б. Бурзин в 1987 г. реконструировал основные этапы формирования планктонной экосистемы (рисунок 22).
РИСУНОК 22. Схема распространения в позднем докембрии и раннем кембрии основных размерных и морфологических групп планктона.
До начала рифея (1,65 млрд лет) фитопланктон состоял почти исключительно из малоразмерных форм с (до 16 мкм). Можно предполагать, что консументы в это время либо были представлены самым мелким размерным классом (назовем его условно "инфузория"), либо отсутствовали вовсе. Впрочем, первое более вероятно: из этого времени известны и мелкие шипастые формы, что может быть первым в истории появлением защитных структур у фитопланктона в ответ на пресс выедания первым зоопланктоном. Для эукариотного фитопланктона, обладающего способностью разрешать "противоречие Виленкина" за счет активных передвижений, выгодно сохранять минимальные размеры тела (что улучшает соотношение поверхность/объем). Поэтому появление в раннем рифее (1,6 млрд лет) форм из следующего размерного класса (16-60 мкм) может рассматриваться как отражение усиления пресса хищников класса "инфузория", и соответственно, попытки фитопланктона уйти за размерный оптимум выедания путем увеличения размера.
Эта стратегия использовалась фитопланктоном на протяжении всего рифея, т.е. почти миллиард лет. В середине рифея (1,4-1,0 млрд лет) появляются акритархи размерного класса 60-200 мкм; это может рассматриваться как реакция на появление консументов следующего размерного класса - "коловратка". К концу же позднего рифея (700 млн лет) фитопланктонные организмы достигают максимального размера за всю историю: помимо доминирующей в это время размерной группы 200-600 мкм в это время появляются, например, гигантские акритархи диаметром до 2-3 миллиметров - это физиологический предел размера для одноклеточных [18]. Доминирование в это время защитной стратегии в виде увеличения размера, при явно подчиненном значении выростов оболочки, свидетельствует о сильном прессе зоопланктона класса "коловратка", но при отсутствии более крупных хищников. Вообще тенденция к однонаправленному изменению размера для жертвы имеет смысл лишь при существовании единственного размерного класса хищников - в противном случае ты тут же попадаешь "из огня в полымя".
А вот дальше произошел вендский фитопланктонный кризис: разнообразие акритарх, достигнув незадолго до появления эдиакарской фауны (600 млн лет) своего максимума, резко падает, причем крупноразмерные формы исчезают полностью. Попытки объяснить этот кризис тем, что фитопланктонные сообщества могли столкнуться в это время с глобальным дефицитом СО2при избытке О2(такое сочетание действительно угнетает развитие микроводорослей), кажутся довольно наивными. Гораздо убедительнее смотрится гипотеза Бурзина о том, что кризис возник в результате появления консументов следующего размерного класса - "циклоп" (до 2 мм [19]), составляющих основную массу современного зоопланктона. От этого врага уже не могла спасти ставшая стандартной стратегия опережающего ухода в следующий размерный класс - ибо фитопланктон уже "уперся" в физиологический предел размеров тела. Лишь к началу кембрия фитопланктону удалось дать адекватный эволюционный ответ на комбинированное воздействие консументов разных классов: с этого момента (и доныне) доминирующей защитной стратегией для фитопланктоных организмов становится образование шипов, выростов, экваториальной каймы и т.п.
Мы уделили столько внимания этой гипотезе по той причине, что рисуемые ею взаимоотношения хищника и жертвы кажутся резко противоречащими современной экологической парадигме, основанной на так называемых циклических балансовых моделях (типа модели Лотки-Вольтерра). Биомассы предыдущего и последующего трофических уровней соотносятся примерно как 10:1 (принцип трофической пирамиды); поэтому ясно, что именно популяция хищника должна подчиняться изменениям в популяции жертвы, но никак не наоборот. Отсюда, как кажется, естественным образом следует и так называемая "филогенетическая реактивность хищника": жертва увеличивает скорость передвижения - хищник вынужден перейти к охоте из засады, жертва одевается панцирем - хищник начинает создавать инструменты для его взлома; именно в таком порядке.
Однако многие палеонтологи (В.А.Красилов, В.В.Жерихин, А.С.Раутиан) приходят в последнее время к выводу о том, что балансовые модели - это путь к познанию функционирования сообществ, но не их эволюции. Все необратимые эволюционные процессы - от глобальных экосистемных кризисов до филогенеза отдельных таксонов - протекают не благодаря, а вопреки сохранению экологического равновесия. Функционирование сообществ основано на циклических процессах, протекающих с отрицательной обратной связью; для того же, чтобы началось развитие, она должна разрушиться и смениться на положительную обратную связь.
Применительно к системе "хищник-жертва" это должно выглядеть так. Любой эволюционный успех жертвы (увеличение скорости передвижения и т.п.) хищник ощущает на своей шкуре немедленно; жертве же вобщем-то наплевать на эволюционные приобретения хищника, поскольку в норме тот все равно изымает лишь небольшую часть ее популяции. До тех пор, пока хищник остается в таком реактивном состоянии, система пребывает в равновесии. Собственно говоря, жертва начинает "замечать" хищника лишь с того момента, как его эффективность превысила некое пороговое значение, в результате чего тот начал подрывать ее ресурсы. В тот самый момент, когда реактивной становится жертва, а не хищник, в системе и возникает положительная обратная связь по типу "гонки вооружений", и система, выйдя из равновесия, начинает эволюционировать в неком (причем совсем не обязательно в "правильном") направлении.
На эволюционную роль верхних трофических уровней экосистемы можно посмотреть и с несколько иной точки зрения. Дело в том, что их взаимодействия с нижними уровнями яляются не столько энергетическими (описываемыми в терминах трофической пирамиды), сколько информационными. Здесь может быть использована аналогия с постиндустриальным обществом, в котором решающее значение приобретает не уровень производства энергии (как это было в индустриальную эпоху), а оптимизация управления энергопотоками за счет информационных связей. Именно таким "блоком управления" экосистемы и являются консументы, деятельность которых вызывает эволюционные изменения у их жертв. Как заметил А.С.Раутиан, само выражение "эволюционные стратегии хищника и жертвы" не вполне правильно: у хищника стратегия есть, а вот у жертвы - одна только тактика [20].
О степени же автономности этого "блока управления" дают наглядное представление именно те палеонтологически документированные ситуации, когда он производит чересчур эффективного хищника, который, по выражению А.Г. Пономаренко, "проедает экосистему насквозь" - как это и произошло при возникновении зоопланктона размерного класса "циклоп". Любой эколог скажет не задумываясь, что "этого не может быть, потому что не может быть никогда!", и будет совершенно прав - но прав именно в рамках своего, экологического, масштаба времени. В эволюционном же масштабе времени такая ситуация (ее можно условно назвать "изобретением абсолютного оружия") не просто возможна - она, как мы увидим далее, служила реальным механизмом запуска целого ряда крупнейших экосистемных перестроек и филогенетических революций.
7. Кембрий: "скелетная революция" и пеллетный транспорт. Эволюция морской экосистемы: кембрий, палеозой и современность. Итак, мы с вами расстались с вендскими экосистемами в момент, когда они пребывали в состоянии глубочайшего глобального кризиса (глава 6). "Эдиакарский эксперимент" окончился неудачей, и все эти загадочные крупноразмерные существа исчезли с лица Земли, не оставив после себя прямых потомков. Фитопланктонный кризис погубил все огромное разнообразие тогдашних акритарх; уцелели лишь немногие, причем морфологически однотипные, формы. В необратимый упадок пришли и бентосные сообщества прокариот-строматолитообразователей; отныне они будут занимать в биосфере подчиненное положение. Хотя тогда появились и некоторые новые группы животных (например, сабелидитиды, строившие органические трубки), конец венда - это время, когда суммарное разнообразие живых организмов на нашей планете падает до рекордно низкого уровня (рисунок 23). И подобно тому, как на развалинах одряхлевшей империи возникает государство молодых энергичных "варваров", на смену докембрийской, прокариотной в своей основе, биосфере стремительно приходит современная, фанерозойская.
Самое же интересное заключается в том, что составляющие эту современную биоту таксоны (а их состав на уровне типов с той поры уже принципиально не менялся!) одновременно обзаводятся минеральным скелетом, причем процесс этот охватывает самые различные группы не только животных, но и водорослей. Вот что пишет, например, один из ведущих специалистов по этому отрезку геологической истории А.Ю. Розанов: "Начнем с того, что поражает любого исследователя, когда он изучает разрезы пограничных отложений докембрия и кембрия. Двигаясь вверх по разрезу от докембрийских пород к кембрийским, мы вдруг обнаруживаем в какой-то момент, что порода насыщена многочисленными и разнообразными остатками организмов, облик которых уже более или менее привычен для нас. Здесь начинают встречаться обычные для всего фанерозоя остатки губок, моллюсков, брахиопод и других организмов. Непривычны только их размеры (первые миллиметры)."
Пытаясь разобраться в причинах этого события, называемого обычно "кембрийской скелетной революцией", надо иметь в виду следующее. Обзавестись твердым скелетом - дело, конечно, "прибыльное", но весьма "капиталоемкое", требующее от организмов больших "первоначальных вложений" по энергии и веществу. Поэтому строить скелет способны лишь существа с достаточно высоким уровнем энергетического обмена, каковой возможен лишь начиная с некоторого уровня содержания на планете свободного кислорода (о "гипотезе кислородного контроля" - см. главу 5). Проанализируем теперь под этим углом зрения докембрийскую экологическую ситуацию.
Начать тут придется издалека. Одной из самых важных задач современной океанологии является детальное изучение морских течений; это, помимо прочего, весьма важно для практики судовождения. С этой целью в воды Мирового океана ежегодно бросают, точно зафиксировав место и время, многие тысячи специальных буев; выловив впоследствии такой буй в другой точке океана, мы получаем представление о направлении и скорости движения переносивших его водных масс. Исследования эти стоят больших денег, а поскольку подавляющее большинство буев при этом безвозвратно теряется, эффективность метода не слишком высока. В связи с этим встал вопрос: а нельзя ли вместо буев найти какой-нибудь естественную "метку" для океанских водных масс?
Такая "метка" действительно существует: это просто-напросто... пыль, которая всегда содержится в атмосфере и, понятное дело, постоянно оседает не только на крышку вашей парты, но и на поверхность океана. Химический состав этой атмосферной пыли в каждой точке Земли свой - на него влияет геохимия соответствующей территории, тип вулканической активности и т.д. А поскольку возможности аналитической химии по нынешнему времени почти безграничны (криминалисты, например, с легкостью определяют, в каком из месторождений мира было добыто контрабандное золото), то для реконструкции картины течений кажется достаточным просто зачерпнуть толику морской воды и установить, в каких именно местах Земли выпадала содержащаяся в воде пыль. Океанологи с энтузиазмом принялись за работу, и спустя небольшое время с изумлением обнаружили, что атмосферной пыли (равно как и вообще взвеси) в океанской воде практически не содержится; на подоконнике, к примеру, пыль будет скапливаться до тех пор, пока ее не сотрет дежурный по классу, а вот из океана она странным образом куда-то улетучивается. Быстро осесть на дно пылинка не может - это можно доказать простыми физическими расчетами. В чем же дело?
К тому времени было уже хорошо известно, какую титаническую работу осуществляют в морях животные-фильтраторы, и прежде всего двустворчатые моллюски; согласно расчетам, одни только уже закартированные устричные банки прогоняют сквозь себя объем воды, равный всему Мировому океану, примерно за месяц! Однако двустворки очищают именно придонные слои воды, куда, как мы помним, пылинкам попасть довольно сложно... Вот тогда-то и выяснилось, что планктонные фильтраторы, и прежде всего - ракообразные, играют в очищении океана роль ничуть не меньшую, чем бентосные. Отсутствие в водах океана сколь-нибудь ощутимых количеств пыли, которая постоянно оседает на его поверхность, является результатом деятельности планктонных ракообразных: они очень быстро отфильтровывают взвесь, упаковывают ее в формируемые их пищеварительным трактом компактные комки отработанной органики - фекальные пеллеты, и отправляют на дно. Без пеллетной транспортировки взвеси мутность воды в океанах была бы несравненно выше.
Именно такая ситуация и должна была наблюдаться в далеком прошлом. Как мы с вами помним, отсутствие в докембрии наземной растительности приводило к многократному усилению эрозии и формированию специфических выположенных ландшафтов - "не суша-не море" (глава 5). Сток с континентов был в основном "плащевым" (т.е. по всей протяженности береговой линии), а равнинные реки и озера, являющиеся ныне главными отстойниками взвеси, отсутствовали. Воды, стекавшие с суши, были мутными, причем самой замутненной в итоге оказывалась именно прибрежная часть океана, наиболее богатая биогенами. Это чрезвычайно затрудняло утилизацию биогенов фитопланктоном из-за узости фотической зоны и резко ограничивало общую продуктивность планктонной экосистемы.
С другой стороны, иной оказывалась и организация самой водной массы. В современных океанах имеется верхний слой воды с резко выраженными сезонными колебаниями температуры - эпиталасса, и нижний, где температура на протяжении всего года практически неизменна - гипоталасса (в озерах им соответствуют эпи- и гиполимнион). На границе между этими слоями располагается узкая зона резкого изменения температуры - термоклин; в современных морях он находится на глубинах от 15 до 100 м. В условиях же замутненной воды энергия солнца практически не проникает глубже первых метров, и термоклин оказывается "подтянутым" под самую поверхность; водная масса оказывается жестко стратифицированой - т.е. разделенную на почти неперемешивающиеся между собою слои: узкую перегретую эпиталассу и относительно холодную, но при этом практически лишенную кислорода гипоталассу. Для этого времени были обычны донные осадки, резко обогащенные неокисленной органикой ("черные сланцы"), которые считают несомненным показателем придонной аноксии (бескислородных условий); позже (начиная с кембрия) их распространение резко сократится. Ситуация аноксии ("заморные обстановки") сильно затрудняла, или даже вообще исключала, существование донной фауны.
И вот на границе венда и кембрия произошло событие, важность которого палеонтологи осознали лишь в самое последнее время (впервые эту идею высказал в 1985 г. А.Г.Пономаренко, а теперь она подтверждена геохимическими данными): в составе докембрийского зоопланктона (бесскелетного, и потому не имевшего шансов захорониться - см. главу 6) возникли достаточно эффективные фильтраторы, формировавшие фекальные пеллеты. Единичные фекальные пелеты были известны и раньше (еще в протерозое), однако именно начало кембрия - это время массового их распространения в морских отложениях.
Следствием этого, вроде бы малозначительного, события стал целый каскад экосистемных перестроек. Уменьшение мутности воды привело к резкому расширению фотической зоны и росту продуктивности экосистем. Расширение же эпиталассы привело к разрушению жесткой температурной стратификации и некоторому обогащению кислородом нижележащих водных слоев. Мало того; мы с вами помним (из главы 6), что именно дисперсная органика, опускающаяся из фотической зоны, является причиной аноксии в придонных слоях - ибо на ее окисление там расходуется кислород; многие исследователи даже употребляют термин "эвтрофный океан". Теперь же органика эта стала, вместе с неорганическими частичками, поступать на морское дно в концентрированном, "упакованном" виде - в пеллетах. Органический детрит, обогащающий донные осадки, - это уже не удушающий эвтрофикатор, а ценный пищевой ресурс; содержание же кислорода в придонных слоях в результате возрастает до уровня, допускающего существование макроскопической донной фауны. Начиная с этого момента в системе возникает положительная обратная связь: придонная фауна в свою очередь начинает очищать воду от органической и минеральной взвеси, количество кислорода в итоге опять возрастает, это позволяет еще дальше наращивать разнообразие живых организмов, и т.д.
Количество свободного кислорода на планете тем временем тоже увеличивается. Во второй половине венда в уже знакомых нам амфибиотических ландшафтах ("ни суша, ни море") место цианобактериальных матов, с их потрясающим по совершенству балансом в производстве и потреблении органики, занимают сообшества водорослей (главным образом - нитчатых), которые иногда называют "водорослевыми болотами". В этих болотах, чрезвычайно широко распространенных во внетропической зоне, захоранивается огромное количество неокисленной органики, а именно такая ситуация (как мы помним из главы 5) вызывает накопление свободного кислорода. По-видимому, сыграло свою роль и крупное Бойканурское оледенение, маркирующее границу венда и кембрия: мы уже помним (см. главу 6), что наличие льдов и психросферы облегчает "закачку" кислорода в глубокие слои океана, вентиляция же дна благоприятствует развитию бентоса.
И вот, наконец, содержание свободного кислорода в океанах достигает той пороговой отметки, начиная с которой становится "экономически оправданным" обзаводиться такой дорогостоящей "высокой технологией", как минеральный скелет. Эта возможность немедленно и одновременно реализуется самыми разными группами животных - от простейших (радиолярии [21] и форамениферы) и губок до членистоногих и моллюсков, - а также растений (известковые водоросли): начинается "скелетная революция" в собственном смысле. Важная деталь: как отмечает А.Ю. Розанов (1986), химический состав этих скелетов, возникающих в самом начале кембрия, настолько разнообразен, что почти полностью исчерпывает все те варианты, которые будут затем возникать на протяжении всего фанерозоя.
Самым лучшим, уникальным, местонахождением кембрийской фауны считают среднекембрийские сланцы Бёджес, открытые в 1912 г. в канадских Скалистых Горах (богатейшие местонахождения в Восточной Сибири, Китае и Гренландии изучены пока недостаточно); тип сохранности в Бёджес таков, что там могут захораниваться и "мягкие" организмы, не имевшие скелета. Так вот, оказалось, что число таких бесскелетных форм в кембрийской фауне даже больше, чем в вендской (хотя доля их теперь невелика). Это служит сильным доводом в пользу того, что в кембрии не появляется вдруг, как по мановению волшебной палочки, некая принципиально новая - "скелетная" - фауна (так иногда полагали прежде), а одевается скелетом та фауна, что уже существует - едва лишь открывается для этого возможность.
Другой неожиданный вывод следует из изучения открытых в конце восьмидесятых годов в Швеции среднекембрийских фосфатных конкреций Ёрстен. При тамошнем типе сохранности фиксируются не покровные ткани (как в Бёждес), а микроскопические щетинки и иные твердые покровные элементы, видимые лишь под сканирующим электронным микроскопом. Оказалось, что в то время существовал многочисленный и очень разнообразный микрозоопланктон - трохофоры (личинки кольчатых червей), зоеа (личинки ракообразных) и т.п. Это позволяет вновь вернуться к гипотезе А.Ю.Розанова (1986) о том, что в кембрии в действительности произошло лишь увеличение размеров уже существовавших на тот момент планктонных организмов, которые по этой причине "выпали в осадок" - перешли к бентическому образу жизни; дополнительные аргументы обретают и построения М.Б. Бурзина (см. выше) о эволюции планктонных экосистем.
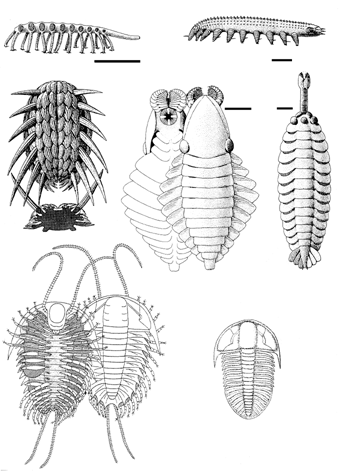
Надо заметить, что мир первых скелетных организмов, сложившийся в начале кембрия (немакит-далдынский ярус), был крайне своеобразен и в чистом виде просуществовал очень недолго, лишь до середины этого периода - хотя отдельные ого представители дожили до середины ордовика. В это время уже существовали достаточно многочисленные животные, имевшие раковины (моллюски и брахиоподы) и жилые трубки (сабелидиты, хиолиты) и т.п.; разнообразнейшие мелкие зубчики - конодонты представляли собою части ротовых органов древнейших вторичноротых - щетинкочелюстных и хордовых. Весьма характерны для этого мира приапулиды (в современных морях эта реликтовая группе насчитывает лишь 20 видов), а также лобоподы (представленные ныне микроскопическими тихоходками и обитающими в подстилке влажных тропических лесов онихофорами); видимо, именно к лобоподам принадлежат многие загадочные кембрийские организмы, вроде галлюциногении (оцените название!). Имелись и и многочисленные артроподоподобные формы (см. ниже) (рисунок 25, а-д).
Основу тогдашних экосистем, однако, составляли водоросли, строившие небольшие известковые постройки - биогермы, и чрезвычайно любопытная, как считалось, вымершая группа организмов - археоциаты. Археоциаты (по-гречески - "древние кубки") представляют собой небольшие прикрепленные ко дну бокалы с двуслойной стенкой (рисунок 24); диаметр их колеблется от нескольких миллиметров до нескольких сантиметров, хотя в Сибири найдены и исполинские формы размером до 1,5 м. Природа их долго вызывала споры (неясно было даже - животные это или растения, или вообще отдельное царство), однако в последнее время почти все исследователи согласны с тем, что их следует относить к губкам. Более того - не так давно в Тихом океане была найдена странная глубоководная губка Vacletia, оказавшаяся по рассмотрении дожившим до наших дней археоциатом и пополнившая собою галерею "живых ископаемых" (вроде кистеперой рыбы или гинкго).
Губки занимают совершенно обособленное положение в системе животного царства и могут быть противопоставлены всем остальным многоклеточным животным: есть веские основания считать, что они ведут свое происхождение от иной группы простейших, нежели прочие метазоа. Существует, например, остроумная и неплохо аргументированная гипотеза Й. Райтнера (1991), согласно которой губки возникают как симбиоз бактериальной колонии с хоанофлагеллятными простейшими [22]. Достигнутый губками уровень организации не позволяет назвать их многоклеточными в строгом смысле ("многотканевыми" по Корлиссу), так что вроде бы нет ничего удивительного в том, что именно эта примитивнейшая группа животных открывает собою фанерозойскую летопись. Но это только на первый взгляд.
Дело в том, что крайне низкий уровень клеточной интеграции оставляет губкам единственно возможную "профессию" - прикрепленный пассивный фильтратор органической взвеси. Любая губка (включая археоциат) представляет собою открытый сверху мешок, стенки которого пронизаны каналами; вода постоянно движется сквозь них во внутреннюю (парагастральную) полость, выходя затем наружу через верхнее отверстие (устье), а входящие в состав стенок каналов хоаноциты (воротничковые жгутиковые клетки) отфильтровывают содержащуюся в воде органику и бактерий. Вопрос для школьной олимпиады: какая сила заставляет воду двигаться сквозь каналы в неподвижной стенке? Ответ: та же самая, что создает печную тягу - разница давлений на высоте поддувала и на высоте конца трубы в соответствии с законом Бернулли [23]: если водная среда, в которой находится губка, имеет ненулевую скорость, то слой движущейся жидкости над устьем создает "подсос". Известное дело: чем выше труба - тем лучше тяга; соответственно, губка может прокачивать сквозь себя воду лишь когда ее устье приподнято над субстратом. (Сходный механизм обеспечивает вентиляцию нор сусликов и иных грызунов. Тот из выходов, через который при рытье норы выбрасывали грунт, оказывается заключен в холмик-"кротовину" и несколько приподнимается над землей; в итоге внутри норы возникает устойчивый ток воздуха от "низкого" выхода к "высокому"). Подъем же устья над субстратом можно обеспечить лишь при наличии твердого опорного скелета - органического или минерального (у современных губок есть оба варианта). Отсутствие в предшествующих отложениях этих легко обнаруживаемых скелетных элементов - спикул - свидельствует о том, что данная группа живых организмов действительно возникла лишь в начале кембрия (возможно, именно по схеме Райтнера - см. выше), т.е. когда уже несомненно существовали и настоящие многоклеточные животные.
Вся геологическая история археоцат (появление, расцвет - около 300 родов, - упадок и вымирание) укладывается в крохотный по геологическим меркам интервал в 15-20 млн. лет - случай совершенно уникальный. Уже во второй половине кембрия эта процветавшая в начале периода группа исчезает "как с белых яблонь дым", и при этом на смену ей не приходит никто - то есть конкурентное вытеснение здесь предположить трудно. Создается впечатление, что археоциаты эксплуатировали некий ресурс, который сперва был в изобилии, а затем стал резко дефицитным. Сопоставляя экологию археоциат (по аналогии с современными губками) и изложенную выше картину вендско-кембрийской биосферной перестройки можно предположить следующее. Группа эта процветала в тот относительно краткий и принципиально неустойчивый период, когда развитие пеллетного транспорта уже обогатило придонные слои кислородом, но еще не сконцентрировало большую часть органики внутри осадка; этот расклад, как полагает А.Г. Пономаренко, должен быть весьма благоприятен для "тонких" фильтраторов. Для того, чтобы эксплуатировать открывшийся ресурс, нет нужды в "квалифицированной рабочей силе" - вполне достаточно макроскопических организмов, даже не являющихся многотканевыми. Однако дальнейшее развитие ситуации с запасанием органики в осадке (что позволяет и сделать ее переработку более равномерной, и улучшить снабжение кислородом придонных слоев) резко ухудшает положение бактерий и пассивных фильтраторов, но зато благоприятствует илоедам. Время археоциат кончается, и наступает время трилобитов (рисунок 25, е-ж) [24].
Давайте теперь вернемся к графику, показывающему изменения разнообразия фауны на протяжении фанерозоя (Рисунок 23). Обсуждаемые нами события знаменуют собой формирование новой биоты, которая сохраняет достаточное единство на протяжении всего палеозоя, и гораздо более разнообразна, чем прежняя, "кембрийская" - и в таксономическом, и в экологическом отношении. В современной экологии достаточнно строго установлено, что смена эвтрофных условий на олиготрофные вызывает в экосистеме более "мелкую нарезку" экологических ниш, и соответственно, рост биологического разнообразия. Все это достаточно хорошо соответствует изложенным выше представлениям об "олиготрофизации" кембрийского океана за счет пеллетного транспорта.
В бентосе тем временем к обитателям раковин и жилых трубок добавляются существа с принципиально иными типами твердого скелета - внутренним (хордовые) и подвижным наружным (членистоногие). Ситуация с членистоногими интересна тем, что они появились позднее многих типов животного царства (во втором веке кембрия - атдабанском), однако сразу в составе практически всех своих подтипов и классов - и ракообразные, и хелицеровые, и вымершие трилобиты, и множество удивительных существ, точная систематическая принадлежность которых до сих пор неясна (рисунок 25, в-д).
Недавно стало выясняться, что ситуация с кембрийскими "членистоногими" (Arthropoda) сходна с той, что была с вендобионтами (см. главу 5): по мере изучения оказывается, что значительная часть из них не может быть отнесена к настоящим артроподам. Например, выяснилось, что у аномалакариса - самого крупного, к слову сказать, животного того времени, достигавшего в размере 70 см - ротовые части состоят из четырех концентрически сходящихся лепестков, образующих нечто вроде зажимов цангового карандаша (рисунок 25, г). Эти лепестки никоим образом не могут представлять собою видоизмененные конечности, что является одним из ключевых диагностических признаков типа членистоногих.
По всей видимости, целый ряд неродственных между собой групп начинает в это время независимо приобретать отдельные признаки членистоногих, и в результате они независимого достигают артроподного уровня организации. Подробнее о процессах параллельной эволюции, подобных этой "артроподизации", мы поговорим позже (в главе 12), пока же для нас существенно лишь само возникновение принципиально нового типа скелета, обладатели которого начинают играть ключевую роль в формирующихся экосистемах. Так, трилобиты становятся, по-видимому, основными потребителями того самого "упакованного" органического детрита, обогащающего донные осадки, и могут считаться настоящей эмблемой палеозоя.
К середине кембрия продукт
|
||||||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 274; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.169 (0.015 с.) |