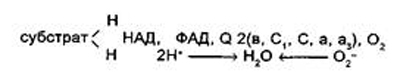Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Современное учение о химизме дыхания. Суть анаэробной фазы дыхания.Содержание книги
Поиск на нашем сайте
Вторая фаза дыхания — аэробная — требует присутствия кислорода. В аэробную фазу вступает пировиноградная кислота. В присутствии достаточного количества О2 пируват полностью окисляется до СО2 и Н2О. Еще в 1910 году шведским химиком Т. Тунбергом было доказано участие органических кислот в процессе дыхания. В 1935 году А. Сент-Дьерди в Венгрии установил, что добавление органических кислот к живой ткани резко активирует поглощение ею кислорода. В 1937 году Г. А. Кребс предложил схему последовательности окисления три- и дикарбоновых кислот до СО2 через «цикл лимонной кислоты» за счет отнятия водорода. Этот цикл и был назван его именем. В цикле окисляется не сама пировиноградная кислота, а ее производное — активный ацетат. Таким образом, первым этапом окислительного расщепления пировиноградной кислоты (ПВК) является процесс образования активного ацетата в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование ПВК осуществляется при участии сложной ферментной системы пируватдегидрогеназы. Эта ферментная система включает в себя три фермента и пять коферментов (тиаминпирофосфат, липовая кислота, коэнзим А, ФАД и НАД). В результате этого процесса образуется активный ацетат (ацетил-КоА), восстановленный НАД (НАДН) и выделяется первая молекула СО2. Ферментная система пируватдегидрогеназа ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение ПВК прекращается. Это один из механизмов регулирования аэробной фазы дыхания. Образовавшийся в ходе этапа активный ацетат вступает в цикл три- и дикарбоновых кислот. Второй этап начинается с взаимодействия активного ацетата с енольной формой щавелево-уксусной кислоты, результатом реакции является образование лимонной кислоты. В этой реакции принимает участие фермент цитратсинтетаза. Следующий этап цикла включает две реакции и катализируется ферментом аконитазой. В первой реакции в результате дегидратации лимонной кислоты образуется цисаконитовая. Во второй реакции цисаконитовая кислота гидратируется в изолимонную. Изолимонная под действием НАД или НАДФ - зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелево-янтарную кислоту, которая декарбоксилируется с образованием оскетоглутаровой кислоты.
Подобно ПВК, α -кетоглутаровая кислота подвергается реакции окислительного декарбоксилирования с аналогичной мультиферментной системой. В ходе этой реакции α -кетоглутаровая кислота превращается в сукцинил-КоА, образуется НАДН и выделяется одна молекула СО2. Подобно активному ацетату, сукцинил-КоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-КоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-КоА может трансформироваться в образовании фосфатной связи АТФ. При участии сукцинил-КоА-синтетазы из сукцинил-КоА, АДФ и Н3РО4 образуется янтарная кислота (сукцинат), АТФ, регенерируется молекула KoA-SH. АТФ образуется в результате субстратного фосфорилирования. На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой, коферментом которой является ФАД. Фумаровая кислота под действием фумаразы, присоединяя воду, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе яблочная кислота с помощью НАД-зависимой малатдегидрогеназы окисляется в щавелево-уксусную (ЩУК). А ЩУК вступает в новый цикл. Следует отметить, что большинство реакций цикла обратимо, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзооргические реакции — цитратсинтетазная и сукцинил-КоА-синтетазная. На протяжении одного оборота цикла при окислении ПВК происходит выделение трех молекул СО2, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль воды в цикле Кребса подтверждает правильность уравнения Палладина, который отмечал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям — коферментов дегидрогеназ) переносится на кислород. Большинство ферментов цикла Кребса локализовано в матриксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрий. Третий этап цикла Кребса — это электрон-транспортная цепь (ЭТЦ). В процессе окисления пировиноградной кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем рассматривать как 2Н+ + 2е-. Именно в таком виде они и передаются по цепи переносчиков. Движущей силой транспорта водорода в дыхательной цепи является разность потенциалов. В связи с этим расположение отдельных переносчиков в дыхательной цепи, также как и в цепи фоосинтетической, определяется величиной их окислительно-восстановительного потенциала. В начале цепи расположен НАД, обладающий наибольшей величиной окислительно-восстановительного (ОВ) потенциала (-0,32В), а в конце — кислород с наиболее положительной величиной (+0,82В).
Остальные переносчики расположены между ними в порядке последовательного повышения положительного потенциала. Это и позволяет электронам передаваться по направлению к кислороду. Б. Чане и др. (США) в 50-х годах предложили следующее расположение переносчиков в ЭТЦ:
Пара электронов от НАДН или сукцината передается по ЭТЦ до кислорода, который, восстанавливаясь и присоединяя два протона, образует воду. Окислительное фосфорилирование. Перенос электронов от НАДН к молекулярному кислороду через ЭТЦ митохондрий сопровождается потерей свободной энергии. В. А. Энгельгардом еще в 30-х годах было показано, что при аэробном дыхании накапливается АТФ. Он первый высказал идею о сопряжении между фосфорилиро-ванным АДФ и аэробным дыханием. В 1937—1939 гг. биохимики В. А. Белицер в СССР и Г. Калькар в США установили, что при окислении промежуточных продуктов цикла Кребса, в частности янтарной и лимонной кислот, образуется АТФ. В анаэробных условиях фосфорилирования не происходит. Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с переносом электронов по ЭТЦ митохондрий, получил название окислительного фосфорилирования. В ЭТЦ синтезируется три молекулы АТФ. Так, первая молекула АТФ образуется при переходе пары электронов от пиридиновой к флавиновой дегидразе. Вторая — при переносе пары электронов от цитохрома б к цитохрому с, и третья — при переносе пары электронов от цитохрома а к цитохромоксидазе (а3). Места образования АТФ в дыхательной цепи не являются случайными. Именно на этих участках выделяется достаточное для образования молекул АТФ количество энергии. Показателем сопряженности окисления и фосфорилирования служит коэффициент фосфорилирования Р/О. В данном случае коэффициент Р/О = 3. Существуют вещества, разобщающие процесс окисления и фосфорилирования (динитрофенол, соли тяжелых металлов). Вся энергия в данном случае превращается в тепло, а коэффициент Р/О = 0. Такое явление может наблюдаться и при засухе или заболевании организма. По поводу механизма окислительного фосфорилирования существует три теории: химическая, конформационная и хемиосмотическая. В настоящее время наибольшим признанием пользуется хемиосмотическая теория английского биохимика Н. Митчелла (1961). Он высказал предположение, что поток электронов через систему молекул-переносчиков сопровождается транспортом ионов Н+ через внутреннюю мембрану митохондрий. В результате на мембране создается электрохимический потенциал ионов Н+, включающий химический или осмотический градиент и электрический градиент (мембранный потенциал). Согласно хемиосмотической теории, электрохимический трансмембранный потенциал ионов Н+ и является источником энергии для синтеза АТФ за счет обращения транспорта ионов Н+ через протонный канал мембранной Н+ АТФ-азы.
Теория Митчелла исходит из того, что переносчики перешнуровывают мембрану, чередуясь таким образом, что в одну сторону возможен перенос и протонов, и электронов, а в другую — только электронов. В результате ионы Н+ накапливаются на одной стороне мембраны. Энергия запасенная, таким образом, используется для синтеза АТФ как результат разрядки мембраны при обратном транспорте протонов через АТФ-азу.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 626; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.93.108 (0.01 с.) |