Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Развитие клеточной теории во второй половине XIX векаСодержание книги
Поиск на нашем сайте
История XVII век 1665 год — английский физик Р. Гук в работе «Микрография» описывает строение пробки, на тонких срезах которой он нашёл правильно расположенные пустоты. Эти пустоты Гук назвал «порами, или клетками». Наличие подобной структуры было известно ему и в некоторых других частях растений. 1670-е годы — итальянский медик и натуралист М. Мальпиги и английский натуралист Н. Грю описали в разных органах растений «мешочки, или пузырьки» и показали широкое распространение у растений клеточного строения. Клетки изображал на своих рисунках голландский микроскопист А. Левенгук. Он же первым открыл мир одноклеточных организмов — описал бактерий и протистов (инфузорий). Исследователи XVII века, показавшие распространённость «клеточного строения» растений, не оценили значение открытия клетки. Они представляли клетки в качестве пустот в непрерывной массе растительных тканей. Грю рассматривал стенки клеток как волокна, поэтому он ввёл термин «ткань», по аналогии с текстильной тканью. Исследования микроскопического строения органов животных носили случайный характер и не дали каких-либо знаний об их клеточном строении. XVIII век В XVIII веке совершаются первые попытки сопоставления микроструктуры клеток растений и животных. К. Ф. Вольф в работе «Теории зарождения» (1759) пытается сравнить развитие микроскопического строения растений и животных. По Вольфу, зародыш как у растений, так и у животных развивается из бесструктурного вещества, в котором движения создают каналы (сосуды) и пустоты (клетки). Фактические данные, приводившиеся Вольфом, были им ошибочно истолкованы и не прибавили новых знаний к тому, что было известно микроскопистам XVII века. Однако его теоретические представления в значительной мере предвосхитили идеи будущей клеточной теории. XIX век В первую четверть XIX века происходит значительное углубление представлений о клеточном строении растений, что связано с существенными улучшениями в конструкции микроскопа (в частности, созданием ахроматических линз). Линк и Молднхоуэр устанавливают наличие у растительных клеток самостоятельных стенок. Выясняется, что клетка есть некая морфологически обособленная структура. В 1831 году Г. Моль доказывает, что даже такие, казалось бы, неклеточные структуры растений, как водоносные трубки, развиваются из клеток. Ф. Мейен в «Фитотомии» (1830) описывает растительные клетки, которые «бывают или одиночными, так что каждая клетка представляет собой особый индивид, как это встречается у водорослей и грибов, или же, образуя более высоко организованные растения, они соединяются в более и менее значительные массы». Мейен подчёркивает самостоятельность обмена веществ каждой клетки. В 1831 году Роберт Броун описывает ядро и высказывает предположение, что оно является постоянной составной частью растительной клетки. Школа Пуркинье В 1801 году Вигиа ввёл понятие о тканях животных, однако он выделял ткани на основании анатомического препарирования и не применял микроскопа. Развитие представлений о микроскопическом строении тканей животных связано прежде всего с исследованиями Пуркинье, основавшего в Бреславле свою школу. Пуркинье и его ученики (особенно следует выделить Г. Валентина) выявили в первом и самом общем виде микроскопическое строение тканей и органов млекопитающих (в том числе и человека). Пуркинье и Валентин сравнивали отдельные клетки растений с частными микроскопическими тканевыми структурами животных, которые Пуркинье чаще всего называл «зёрнышками» (для некоторых животных структур в его школе применялся термин «клетка»). В 1837 году Пуркинье выступил в Праге с серией докладов. В них он сообщил о своих наблюдениях над строением желудочных желёз, нервной системы и т. д. В таблице, приложенной к его докладу, были даны ясные изображения некоторых клеток животных тканей. Тем не менее установить гомологию клеток растений и клеток животных Пуркинье не смог:
Сопоставление клеток растений и «зёрнышек» животных Пуркинье вёл в плане аналогии, а не гомологии этих структур (понимая термины «аналогия» и «гомология» в современном смысле). Школа Мюллера и работа Шванна. Второй школой, где изучали микроскопическое строение животных тканей, была лаборатория Иоганнеса Мюллера в Берлине. Мюллер изучал микроскопическое строение спинной струны (хорды); его ученик Генле опубликовал исследование о кишечном эпителии, в котором он дал описание различных его видов и их клеточного строения. Здесь были выполнены классические исследования Теодора Шванна, заложившие основание клеточной теории. На работу Шванна оказала сильное влияние школа Пуркинье и Генле. Шванн нашёл правильный принцип сравнения клеток растений и элементарных микроскопических структур животных. Шванн смог установить гомологию и доказать соответствие в строении и росте элементарных микроскопических структур растений и животных. На значение ядра в клетке Шванна натолкнули исследования Матиаса Шлейдена, у которого в 1838 году вышла работа «Материалы по фитогенезу». Поэтому Шлейдена часто называют соавтором клеточной теории. Основная идея клеточной теории — соответствие клеток растений и элементарных структур животных — была чужда Шлейдену. Он сформулировал теорию новообразования клеток из бесструктурного вещества, согласно которой сначала из мельчайшей зернистости конденсируется ядрышко, вокруг него образуется ядро, являющееся образователем клетки (цитобластом). Однако эта теория опиралась на неверные факты. В 1838 году Шванн публикует 3 предварительных сообщения, а в 1839 году появляется его классическое сочинение «Микроскопические исследования о соответствии в структуре и росте животных и растений», в самом заглавии которого выражена основная мысль клеточной теории:
XX век Клеточная теория со второй половины XIX века приобретала всё более метафизический характер, усиленный «Целлюлярной физиологией» Ферворна, рассматривавшего любой физиологический процесс, протекающий в организме, как простую сумму физиологических проявлений отдельных клеток. В завершении этой линии развития клеточной теории появилась механистическая теория «клеточного государства», в качестве сторонника которой выступал в том числе и Геккель. Согласно данной теории организм сравнивается с государством, а его клетки — с гражданами. Подобная теория противоречила принципу целостности организма. Механистическое направление в развитии клеточной теории подверглось острой критике. В 1860 году с критикой представления Вирхова о клетке выступил И. М. Сеченов. Позднее клеточная теория подверглась критическим оценкам со стороны других авторов. Наиболее серьёзные и принципиальные возражения были сделаны Гертвигом, А. Г. Гурвичем (1904), М. Гейденгайном (1907), Добеллом (1911). С обширной критикой клеточного учения выступил чешский гистолог Студничка (1929, 1934). В 1930-х годах советский биолог О. Б. Лепешинская, основываясь на данных своих исследований, выдвинула «новую клеточную теорию» в противовес «вирховианству». В её основу было положено представление, что в онтогенезе клетки могут развиваться из некоего неклеточного живого вещества. Критическая проверка фактов, положенных О. Б. Лепешинской и её приверженцами в основу выдвигаемой ею теории, не подтвердила данных о развитии клеточных ядер из безъядерного «живого вещества». Электронная микроскопия
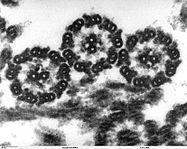
В 30-х годах XX века был сконструирован электронный микроскоп, в котором вместо света через объект пропускается пучок электронов. Теоретический предел разрешения для современных электронных микроскопов составляет около 0,002 нм, однако из практических причин для биологических объектов достигается разрешение только около 2 нм. С помощью электронного микроскопа можно изучать ультраструктуру клеток. Различают два основных типа электронной микроскопии: сканирующую и трансмиссионную. Сканирующая (растровая) электронная микроскопия (РЭМ) используется для изучения поверхности объекта. Образцы зачастую покрывают тонкой пленкой золота. РЭМ позволяет получать объемные изображения. Трансмиссионная (просвечивающая) электронная микроскопия (ПЭМ) — используется для изучения внутреннего строения клетки. Пучок электронов пропускается через объект, предварительно обработанный тяжелыми металлами, которые накапливаются в определенных структурах, увеличивая их электронную плотность. Электроны рассеиваются на участках клетки с большей электронной плотностью, в результате чего на изображениях эти области выглядят темнее
Метод центрифугирования
Его используют, когда необходимо отделить мембраны клеток от общей структуры; метод меченых атомов. Применяют для изучения биохимических процессов в клетках: для этого в них вводят меченый радиоактивный изотоп; прижизненное изучение. Этот метод исследования позволяет изучить динамические процессы, происходящие в клетке. Заключение цитологического исследования основывается на особенностях изменения цитоплазмы, ядра клетки, ядерно-цитоплазменного соотношения, образования комплексов и структур клеток. Применяют цитологический анализ при профилактическом осмотре, для уточнения диагноза, во время операции, для своевременного выявления рецидивов, контроля над ходом лечения.
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
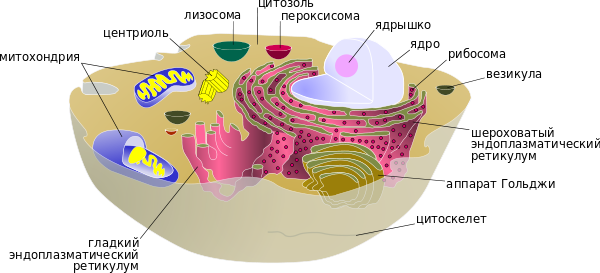
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам. Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом. Прокариотическая клетка Прокариоты (от лат. pro — перед, до и греч. κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма. Эукариотическая клетка Эукариоты (эвкариоты) (от греч. ευ — хорошо, полностью и κάρῠον — ядро, орех) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды. Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре, характерными их признаками являются отсутствие ядра и мембранных органелл. Основными компонентами прокариотической клетки являются:
Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов.
Одномембранные структуры: Комплекс Гольджи Вакуоли Двухмембранные структуры: Ядро Хромосомы Митохондрии Пластиды Немембранные структуры: Ложноножки Реснички и жгутики Базальные тельца Рибосомы Клеточный центр Включения Структура цитоплазмы Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
На фотографиях зелёный флуоресцентный белок показывает расположение различных частей клетки Аппарат Гольджи Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны располагающиеся ближе к ядру клетки (цис -Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы (транс -Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки. Ядро Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Лизосомы Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной. В ней находятся литические ферменты, способные расщепить все биополимеры. Основная функция — автолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки. Цитоскелет К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка. Центриоли Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3. Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки. Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей. Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек. Митохондрии Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий. Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки, так называемые кристы. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии. Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что, безусловно, указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов. Самое большое число митохондриальных генов (97) из изученных организмов имеет простейшее Reclinomonas americana.
*Константа седиментации (скорость оседания в ультрацентрифуге)
Клеточный цикл
Амито́з — прямое деление клетки, происходит в соматических клетках эукариот реже, чем митоз. В большинстве случаев амитоз наблюдается в клетках со сниженной митотической активностью: это стареющие или патологически измененные клетки, часто обреченные на гибель (клетки зародышевых оболочек млекопитающих, опухолевые клетки и другие). При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует. Спирализация хроматина не происходит, хромосомы не выявляются. Клетка сохраняет свойственную ей функциональную активность, которая почти полностью исчезает при митозе. Таково, например, деление макронуклеусов многих инфузорий, где без образования веретена происходит сегрегация коротких фрагментов хромосом. При амитозе делится только ядро, причём без образования веретена деления, поэтому наследственный материал распределяется случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в дальнейшем не способны вступать в нормальный митотический цикл. При повторных амитозах могут образовываться многоядерные клетки. Мито́з (от греч. μιτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток, один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяции тканевых клеток. Биологическое значение митоза заключается в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений. На основании морфологических особенностей митоз условно подразделяется на:
Продолжительность митоза в среднем составляет 1—2 часа. В клетках животных митоз, как правило, длится 30—60 минут, а в растительных — 2—3 часа. Клетки человека за 70 лет суммарно претерпевают порядка 1014 клеточных делений.
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять — профаза, прометафаза (метакинез), метафаза, анафаза и телофаза (или цитотомия) — составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы. Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза (2—270 минут) и телофаза (1,5—140 минут). Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза (0,3—175 минут) и анафаза (0,3—122 минуты). Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.
К рисунку. Деление растительной клетки с крупной центральной вакуолью:
К рисунку. Начальные стадии митоза растительной клетки: Препрофаза Препрофаза — редко используемый термин для обозначения дополнительной стадии митоза растительной клетки. К основным событиям препрофазы можно отнести: оформление препрофазного кольца, образование фрагмосомы и начало нуклеации микротрубочек вокруг клеточного ядра. Несмотря на наличие термина «препрофаза» перечисленные события чаще рассматриваются в составе фазы G2 или в составе профазы. В клетках богатых вакуолями в ходе препрофазы образуется фрагмосома — одна из структур, определяющих плоскость деления растительной клетки. Фрагмосома представляет собой слой цитоплазмы, пересекающий вакуоль в плоскости клеточного деления. Ядро в клетках с крупной центральной вакуолью обычно располагается на периферии. Во время препрофазы оно перемещается в область фрагмосомы. В ходе перемещения ядра вакуоль рассекается полосками цитоплазмы, содержащими элементы цитоскелета. Во фрагмосоме также образуется митотическое веретено. Во время цитокинеза в области фрагмосомы формируется фрагмопласт и новая клеточная стенка. Одновременно с фрагмосомой образуется препрофазное кольцо, и обе структуры располагаются в одной плоскости. Препрофазное кольцо — это кольцеобразное скопление микротрубочек и актиновых филаментов вблизи клеточной мембраны в плоскости деления растительной клетки. Ядро располагается в центре препрофазного кольца и связано с ним радиально расходящимися микротрубочками. Внешне данная структура напоминает колесо с ободом и спицами из микротрубочек и актиновых филаментов, а также с ядром на месте втулки. Структура кольца также обогащена элементами ЭПР и везикулами аппарата Гольджи. Препрофазное кольцо формируется перед профазой митоза. После начала профазы микротрубочки кольца деполимеризуются и далее участвуют в образовании веретена деления. Функции препрофазного кольца пока не ясны. Однако подмечено, что цитокинез растительной клетки происходит в плоскости определённой положением препрофазного кольца. При симметричном делении кольцо формируется в середине, а при асимметрическом делении образуется ближе к одному концу клетки. Профаза К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью. Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца (центросомы), микротрубочки и кинетохоры хромосом. С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут (в интерфазе) до 1,5 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретен
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-27; просмотров: 234; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.25.130 (0.012 с.) |