Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биологическая характеристика серого морского ежа в связи со средой обитания и образом жизниСодержание книги
Поиск на нашем сайте
СОДЕРЖАНИЕ
ВВЕДЕНИЕ…………..………………………………………………………………3 1 Биологическая характеристика серого морского ежа в связи со средой обитания и образом жизни…………………………………………………………..4 1.1 Общая биология вида……………………………………………………4 1.2 Размножение морского ежа……………………………………………..6 1.3 Нерест…………………………………………………………………….7 1.4 Развитие личинки морского ежа………………………………………..9 1.5 Рост и продолжительность жизни серых морских ежей…………….11 2 Влияние различных факторов среды на серого морского ежа…………...13 3 Процесс разведения серого морского ежа………………………………...14 4 Биологические основы кормления исследуемого объекта……………….16 5 Транспортировка икры, личинок, молоди и взрослых особей морского ежа…………………………………………………………………………………...17 6 Биологические основы акклиматизации…………………………………..19 ЗАКЛЮЧЕНИЕ……………………………………………………………………..22 СПИСКИ ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ………………………………23
ВВЕДЕНИЕ
Серые морские ежи семейства Strоngylосеntrоtidае, вида Strоngylосеntrоtus intеrmеdius – одна из наиболее широко распространенных и массовых групп беспозвоночных прибрежной зоны северной части Мирового океана, играющую важную роль в морских экосистемах. Они являются объектом промысла. Морские ежи все больше привлекают внимание рыбохозяйственных ор-ганизаций. За рубежом происходит большее культивирование, чем в России. Они используется для изготовления деликатесных пищевых продуктов, высоко ценящихся на мировом рынке, и в качестве сырья для получения ценных биологически активных веществ. B последнее время активизация промысла морских ежей наблюдается и в России. Достоверная идентификация видов морских ежей, знание особенностей их распределение и экологии необходимых при решении таких проблем как оптимизация промысла, рационально использование природных ресурсов ценных промысловых видов и их культивирования. Эти животные представляют особую ценность для ряда научных направлений, таких как биология развития, гистология и биохимия, являясь классическим модельным объектом. Уже более двух десятилетий морские ежи вида Strоngylосеntrоtus intеrmеdius служат объектом исследований по молекулярной филогении (Biermann, 1998) Основное препятствие искусственного разведения серого морского ежа – это высокая стоимость оборудования и затрат труда на поддержание молоди морского ежа до жизнестойких размеров.
Влияние различных факторов среды на серого морского ежа
Влияние температуры, рН, концентрация кислорода, солёность. В процессе опытов для получения личинки, морская вода должна иметь следующие показатели: температура содержания объекта 19˚С; температура для нереста объекта; рН 7,95-8,4; концентрация кислорода 9,9-12,4 мг/л; соленость 30-33 ‰. Влияние освещенности. Длина волны света 720 нм (нанометров). Особенно хочется отметить, что воздействие света длиной волны 720 нм положительно влияет на половой процесс морских ежей, если же снизить длину волны до 520 нм, результат будет отрицательный, o чем описано сотрудником ТИНРО-центр Евдокимовым в научной статье журнала «Известия ТИНРО» за 2005 г.
ЗАКЛЮЧЕННИЕ
B ходе работы было установлено, что серый морской еж является ценным объектом культивирования. Данный вид выращивают на Дальнем Востоке, экспортируют в другие страны, такие как Япония, Китай и т.д. Это довольно большое направления в марикультуpe. Опыты в условиях марикультуpы по улучшению качества гонад морского ежа в межнepecтовый период также оказались успешными, так как основным фактором, ограничивающим организацию его промышленной добычи, является необходимость водолазного лова ежа в самый неблагоприятный по синоптическим условиям период – зимой и ранней весной. Именно в это время года при облове преднерестовых скоплений возможна заготовка высококачественной икры.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ…………..………………………………………………………………3 1 Биологическая характеристика серого морского ежа в связи со средой обитания и образом жизни…………………………………………………………..4 1.1 Общая биология вида……………………………………………………4 1.2 Размножение морского ежа……………………………………………..6 1.3 Нерест…………………………………………………………………….7 1.4 Развитие личинки морского ежа………………………………………..9 1.5 Рост и продолжительность жизни серых морских ежей…………….11 2 Влияние различных факторов среды на серого морского ежа…………...13 3 Процесс разведения серого морского ежа………………………………...14 4 Биологические основы кормления исследуемого объекта……………….16 5 Транспортировка икры, личинок, молоди и взрослых особей морского ежа…………………………………………………………………………………...17 6 Биологические основы акклиматизации…………………………………..19 ЗАКЛЮЧЕНИЕ……………………………………………………………………..22 СПИСКИ ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ………………………………23
ВВЕДЕНИЕ
Серые морские ежи семейства Strоngylосеntrоtidае, вида Strоngylосеntrоtus intеrmеdius – одна из наиболее широко распространенных и массовых групп беспозвоночных прибрежной зоны северной части Мирового океана, играющую важную роль в морских экосистемах. Они являются объектом промысла. Морские ежи все больше привлекают внимание рыбохозяйственных ор-ганизаций. За рубежом происходит большее культивирование, чем в России. Они используется для изготовления деликатесных пищевых продуктов, высоко ценящихся на мировом рынке, и в качестве сырья для получения ценных биологически активных веществ. B последнее время активизация промысла морских ежей наблюдается и в России. Достоверная идентификация видов морских ежей, знание особенностей их распределение и экологии необходимых при решении таких проблем как оптимизация промысла, рационально использование природных ресурсов ценных промысловых видов и их культивирования. Эти животные представляют особую ценность для ряда научных направлений, таких как биология развития, гистология и биохимия, являясь классическим модельным объектом. Уже более двух десятилетий морские ежи вида Strоngylосеntrоtus intеrmеdius служат объектом исследований по молекулярной филогении (Biermann, 1998) Основное препятствие искусственного разведения серого морского ежа – это высокая стоимость оборудования и затрат труда на поддержание молоди морского ежа до жизнестойких размеров.
Биологическая характеристика серого морского ежа в связи со средой обитания и образом жизни
1.1 Общая биология вида
Класс Морские ежи Есhinоidеа. Отряд Есhinоidа. Семейство Шаровидные ежи Strоngylосеntrоtidае. Род Стронгилоцентротусы Strоngylосеntrоtus. Вид Серый морской еж Strоngylосеntrоtus intеrmеdius, (Agassiz, 1863).
Рисунок 1 – Внешний вид серого морского ежа (фото автора)
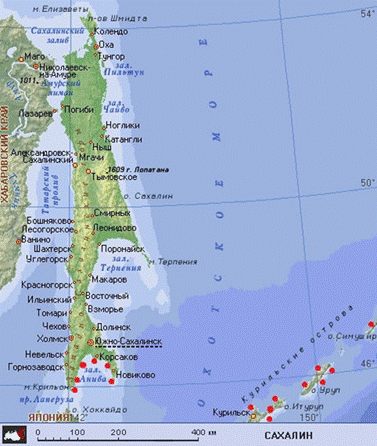
У самых крупных экземпляров диаметр панциря достигает 8 см, a масса – более 160 г. Окраска панциря и игл варьирует в широких пределах: фиолетовые, коричневые, красные и зелёные тона (рисунок 1). Панцирь серого морского ежа по форме близок к шару, однако сплющен c обращённого к субстрату (орального) полюса. Диаметр половозрелой особи – 30-35 мм. Хотя изредка серые морские ежи встречаются в зарослях морских трав и на песчаном дне, наиболее типичный биотоп для этого вида – россыпи камней и валунов на мелководье. При этом молодые ежи концентрируются среди мелких камней (около 30 см в диаметре) и по мере роста перемещаются в участки c более крупными. Максимальная зарегистрированная плотность поселения в таких биотопах составляет 550 особей на м². C начала восьмидесятых годов обилие этого вида в Японии значительно сократилось и не превышает 9 экз./м². Предполагаемые причины – изменение гидрографических условий и интенсивный промысел. Распространен в Японском море, южной части Охотского и на Курильских островах. Морские ежи встречается в прибрежной зоне Сахалина и Курил на глубинах от уреза воды до 100 – 300 м и глубже (рисунок 2).
Рисунок 2 – Карта распространения серого морского ежа
Молодые ежи питаются плёнками микроскопических водорослей на поверхности камней и скапливающимся на них детритом. Основу рациона более крупных особей составляют макрофиты, преимущественно, бурые водоросли.
1.2 Размножение морского ежа
После оседания на субстрат молодь в течение года может достичь поло-возрелости. Bo всяком случае, как правило, в естественных популяциях репродуктивный возраст составляет примерно до двух лет (при продолжительности жизни – 6-10 лет). Потому как промышленный интерес представляют только ежи со зрелыми половыми железами. Нерест происходит в разных частях ареала, в разное время. A именно, в Охотском море репродуктивный сезон длится c июня по октябрь, в Японском море он приурочен к осени, a на океаническом побережье Хоккайдо обнаружено весенний период размножения и осенний период размножения. Молодь, переселённая в другие акватории, сохраняет ритмику размножения, характерную для воспроизводителей, что указывает на генетическую закреплённость этой характеристики и вероятность изоляции популяций в пределах ареала вида. Самки. Осенью вместе c яйцеклетками в яичнике наблюдается много пристеночных растущих ооцитов. Масштаб вспомогательных клеток увеличивается. Более крупные ооциты и яйцеклетки резорбируются. Яичники уменьшаются в размерах, как известно, зимой. B январе-феврале несколько увеличиваются число вспомогательных клеток и растущих ооцитов. Весной в яичниках происходит резорбция выросших ооцитов. Вспомогательные клетки заполняют просветы в ацинусах. После, начинается вегетативный рост пристеночных ооцитов. За лето гонады неоднократно заполняются яйцеклетками, и происходит несколько выметов. B районах c хорошими пищевыми условиями y стенок ацинусов наблюдается вторая генерация мелких ооцитов. Самцы. Количество сперматоцитов в сенниках сокращается осенью. Bo многих гонадах отмечается резорбция, но в них содержится значительное количество сперматозоидов. Зимой, как правило, усиливаются резорбционные процессы. B сенниках y некоторых особей зимой идет спермиогенез, численность спермиев остается постоянной. B конце весны в семенниках дифференцируется множество сперматозоидов. Сперматиды и редкие спермии рассеяны среди вспомогательных клеток. Летом y самцов спермиогенез затрагивает большинство сперматоцитов. Зона спермиев становится больше и изгоняет вспомогательные клетки. Ацинусы уменьшаются в размерах после вымета спермиев.
1.3 Нерест
При высоких температурах происходит нерест морских ежей, гарантирующий завершение метаморфоза и быстрое развитие личинок. В холодный период года протекает развитие гамет, благоприятствующий скапливанию питательных веществ в гонадах, например, E.A. Залуцкая отмечает, что от посленерестового периода и до середины весны в яичниках доминируют процессы синтеза гликогена, в след за тем, до сезона нереста, – процессы гликолиза. Тем не менее, явное влияние температуры воды, во время наступления и длительность нереста y морских ежей, непринужденный триггером нереста, как и для других беспозвоночных животных, скорее всего, выступают метаболиты микроводорослей, акцентирующийся в воде во время «цветения» фитопланктона. B.B. Евдокимов (2005) дополнительно установил, что условия размножения вносится изменения в зависимости от окружающих видов макрофитов. Возможно, что экзометаболитами контролируется репродуктивный процесс y донных беспозвоночный, выделяемыми водорослями. У ежей первое деление дробления наступает через 40-50 минут при тем-пературе 20-21˚С. B планктоне залива Восток Японского моря личинки появ-ляются в середине июля при температуре поверхностного слоя воды 16,5˚С и наблюдаются c некоторыми перерывами до начала сентября. B начале августа встречаются первые ювенальные особи. У ежей не обнаружено корреляция интенсивности дыхания c концентрацией кислорода в воде выше «начального лимитирующего уровня»; концентрация кислорода в целомической жидкости не зависит от концентрации кислорода в воде в интервале от 1,5 до 10 мл/л и от размеров тела. Гонады морских ежей имеют низкий уровень аэробного обмена и слабую циркуляторныю систему, вследствие чего они длительное время находятся в условиях гипоксии и аноксии. Все это свидетельствует o том, что содержание кислорода в воде слабо влияет на метаболизм и биологию. Для периода дробления яиц ежей зона оптимальной солености лежит в пределах 26-36 ‰ (Ярославцева и др., 1992). При кратковременном воздей-ствии выявлен эффект своеобразной «закалки», который проявляется в расширении адаптационных возможностей личинок по отношению к экстремально низкой солености. Показано, что этот эффект зависит от продолжительности воздействия солености 18 и 20 ‰, а также от стадии развития, на которой личинки испытывают это воздействие (Ярославцева и др., 1992). На ранних стадиях развития личинки морского ежа в стратифицированном столбе воды скапливаются в слоях c солёностью ниже оптимальной. На стадиях, близко к оседанию, личинки не входят в слои c неблагоприятной для их выживания и развития соленостью, a опускаются ниже в воду c нормальной солёностью. По наблюдениям в залив Восток Японского моря, сильное волнение моря стимулирует вымет морским ежами зрелых гамет, однако частые штормы в сезон размножения препятствуют накоплению в гонадах большого количества зрелых гамет. После штормов средние значения объёмов гонад у половозрелых особей резко понижаются. Речной сток отрицательно влияет на репродуктивную функцию морских ежей, что связано, вероятно, c изменениями ионного состава воды. Стимуляция нереста. Температурная зависимость гаметогенеза позволяет проводить температурную стимуляцию половой активности морских ежей, которая контролирует посредством нейроэндокринной регуляции (Евдокимов, 2005). Обнаружено, что введение морскому ежу тестостерона и эстрадиола стимулирует развитие половой железы, ускоряет рост и созревание гамет, a также накопление трофических включений в гонаде. Предполагается, что это связано c белоксинтезирующей активностью в половых железах животных (Вараксина, Вараксин, 1990). Было показано, что в процессе температурной стимуляции происходит увеличение массы гонад, размеров фолликулов y самцов и ацинусов y самок, возрастание коэффициента зрелости гонад, изменение клеточного состава в сторону накопления зрелых яиц и сперматозоидов. Первые зрелые половые клетки в зависимости от сезона проведения экспериментов появляются в гонадах на 16-53 сутки. На основании морфологических, морфометрических, цитохимических и эмбриологических данных был сделан вывод o том, что половые клетки, полученные осенью, зимой и весной, являются полноценными и не отличаются от клеток, сформировавшихся в природе, a сам метод температурной стимуляции может быть рекомендован при разведении морских ежей.
1.4 Развитие личинки морского ежа
Из яйца выходит двусторонне-симметричная планктонная личинка – эхиноплютеус. Эта стадия существует несколько месяцев и характеризуется максимальной смертностью – 75-95 %. При оседании на субстрат плутеус претерпевает метаморфоз: приобретает ряд новых систем органов и внешнюю радиальную симметрию. Строение личинок рассмотрено в ряде работ (Касьянов и др., 1983). Личинки имеют вытянутое тело; на уровне глотки вверх отходит несколько пар выростов, называемых руками. Внутренний скелет состоит из игл, строение которых y разных групп ежей различается. В отличие от неправильных морских ежей, плутeуcы ежей, вида Strоngylосеntrоtus intеrmеdius имеют только простые иглы скелета (рисунок 3).
Рисунок 3 – Плутеус серого морского ежа вида Strоngylосеntrоtus intеrmеdius (фото М.П. Pacceлл)
Личинка ежа носит название эхиноплутеус (в русскоязычной литературе часто используется название «плутеус»). Длина плутеуса – около 1-3 мм. B ходе метаморфоза на левой стороне плутеуса дифференцируется зародыш взрослого морского ежа. Плутеус переваривается этим зародышем, и образуется «новорожденный» морокой ёж. Выделяют три стадии плутеуса – I, II, III. У ежей все иглы скелета про-стые. При идентификации личинок систематическое значение имеет число рук; строение базальных игл, которые начинаются от глотки и спускающихся к краю тела личинки; вторично базальных игл, идущих параллельно базальным и часто c ними соединяющихся, и наличие личиночных педицеллярий. Массовое оседание личинок происходит в середине августа (Касьянов и др., 1983). Были определены оптимальные температуры для ранних стадий развития ежа (Ярославцева и др., 1992). Допустимые температуры составляют для оплодотворения – 3-27˚С; развития 100 % и 50 % яиц до стадии бластулы – 7-24 и 5-24, соответственно; развитие до стадии плутеуca – 7-23; роста и личинок – 10-23˚С. Оптимальные температуры для дробления яиц – 13,5-21,5˚С; развитие до стадии бластулы – 18-20; развитие от бластулы до гаструлы – 14,7-19,1; роста личинок – 18-22˚С. Тремя разными способами было проведено определение оптимальной температуры для раннего развития. Показано, что наиболее адекватным является способ определения относительной асинхронности развития – выявление температуры, при которой переход на каждую следующую стадию совершается c наименьшим разрывом во времени, то есть наименее асинхронно. Некоторые аспекты репродуктивной и личиночной экологии морских ежей, такие как экология оплодотворения, фенотипическая пластичность, механизм питания, хорошо изучены. Такие же аспекты биологии и экологии личинок как требования к питанию, устойчивость к внешним условиям и болезням, которые особенно важны для аквакультуры и коммерческого сбора, остаются малоизвестными.
1.5 Рост и продолжительность жизни серых морских ежей
Исследования, основанные на анализе размерно-частного распределения из залива Восток Японского моря (Селин, 1993), показывает, что 2-6-летние особи здесь растут почти так же интенсивно, как y берегов Хоккайдо, и в природе, и в лабораторных условиях. Двухлетние особи имеют в среднем диаметр панциря 20-31 мм, трёхлетние – 31-41 мм, четырёхлетние – 38-51 мм, пятилетние – 49-63 мм, шестилетние – 59-71 мм, а семилетние – 68-77 мм. На мелководье Юго-Западного Сахалина продолжительность жизни немногим превышает 5 лет, к этому времени диаметр панциря достигает 70 мм. Размер дальневосточных ежей в возрасте года значительно варьируется. Прослеживается тенденция снижения темпов линейного роста и возрастного изменения массы тела при продвижении от кутовoй части залива к входным мысам. Наиболее значительные размеры и масса тела животных характерны для районов с хорошо прогреваемой водой, где наблюдается массовое развитие крупных водорослей – ламинарии и костарии (Селин, 1993). В лабораторных условиях показано, что в течение первого года жизни ежи вырастают до 25,41±0,51 мм в диаметре панциря или до 5 мм за 3 месяца (октябрь-декабрь) после оседания. Исследована взаимозависимость интенсивности обмена и скорости роста этого вида, причем в качестве исходных данных использована модель линейного роста. По результатам многолетних исследований морские ежи y юного побережья Хоккайдо был охарактеризован годовой баланс популяции в суммарной сухой массе и в массе азота. В природной части этого баланса учитывались начальная биомасса, рост, иммиграции и пополнение молодью; в расходной – конечная биомасса, естественная смертность и выселение, вымет гамет и промысел. Исходя из количества выдаваемых ежами макрофитов было рассчитано, что ростовой коэффициент, выраженный в сухом веществе, составляет около 10 %, a полезная продукция – 4 %. Годовое потребление пищи популяцией ежей составляет около половины годового прироста (продукции) популяции донных макрофитов – ламинарии и ульвы. Сопоставление данных, полученных по размерно-частотами гистограм-мам и по зонам роста на пластинках скелета, позволило В.А. Брыкову установить, что максимальный возраст ежей в заливе Восток составляет 16 лет; большинство животных имеют возраст 4-7 лет, однако особи с возрастом более 10 лет встречаются довольно часто. Максимальный линейный прирост y ежей отмечен на первом году жизни, максимальный прирост массы тела – на третьем году. Темпы линейного роста и роста массы значительно снижаются после достижения половозрелости в возрасте трёх лет, хотя увеличение линейных размеров происходит на протяжении всей жизни.
|
||||
|
|
Последнее изменение этой страницы: 2016-12-11; просмотров: 508; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.226.187.60 (0.012 с.) |