Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Высокоспецифичные протеиназыСодержание книги Поиск на нашем сайте
Адсорбционный центр этих ферментов имеет сложное строение. Они способны распознавать структуру радикала не только одной из аминокислот, но целого участка полипептидной цепи, в составе которого находится пептидная связь, которую гидролизует фермент. Часто высокоспецифичная протеиназа может узнать и гидролизовать только одну связь из сотен других, имеющихся в белке-субстрате. Такое высокоспецифичное расщепление молекулы белка в одном строго определенном месте называется “ограниченный протеолиз”. Высокоспецифичные протеиназы можно разделить на две группы: 1. Внутриклеточные высокоспецифичные протеиназы. Обеспечивают постсинтетическую модификацию белка. В результате реакций ограниченного протеолиза полипептидная цепь укорачивается и белок приобретает биологическую активность. К внеклеточным протеиназам относится система свертывания крови, система комплемента и система регуляции сосудистого тонуса. Общая характеристика протеосом Другая хорошо регулируемая система деградации белков локализована в цитоплазме. Она состоит из больших белковых комплексов (молекулярная масса 2 ּ 106 Да), протеосом. Протеосомы содержат бочковидное ядро из 28 субъединиц. Протеолитическая активность (на рис.2 показана в виде ножниц) локализована во внутреннем 20S-ядре. С торцов бочки запираются сложно устроенными 19S-частицами, контролирующими доступ в ядро. Белки, которым предстоит разрушение в протеасоме (например, содержащие ошибки транскрипции или состарившиеся молекулы), метятся путем ковалентного связывания с небольшим белком убиквитином. Убиквитин активирован благодаря наличию тиолсложноэфирной связи. Меченые убиквитином («убиквитинированные») молекулы распознаются 19S-частицами с потреблением АТФ и попадают в ядро, где происходит их деградация. Убиквитин не разрушается и после активации используется вновь.
КАТАБОЛИЗМ АМИНОКИСЛОТ. 80% аминокислот, которые поступают в организм из желудочно-кишечного тракта, используются для синтеза белков. Остальные 20% вступают в метаболические процессы. Все эти процессы можно разделить на 2 группы: 1. Общие пути катаболизма аминокислот (для всех аминокислот они одинаковы). В них принимает участие общая часть молекулы аминокислоты.
2. Специфические пути метаболизма для каждой отдельной аминокислоты (разные для разных аминокислот) - участвуют радикалы аминокислот. Это - особенности обмена отдельных аминокислот. ОБЩИЕ ПУТИ КАТАБОЛИЗМА АМИНОКИСЛОТ 1. Декарбоксилирование 2. Дезаминирование 3. Трансаминирование (переаминирование)
ДЕКАРБОКСИЛИРОВАНИЕ
В организме человека происходит только окислительное декарбоксилирование. Ферменты - декарбоксилазы. Их простетическая группа представлена пиридоксальфосфатом - это активная форма витамина В6. Продукты реакции – биогенные амины и углекислый газ. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕКАРБОКСИЛИРОВАНИЯ АМИНОКИСЛОТ 1. Реакции необратимы - приводят к необратимому распаду аминокислот. 2. Образуется значительное количество СО2 - конечного продукта метаболизма, который выводится из организма. 3. Образуются амины, которые обладают высокой биологической активностью. Поэтому такие амины называют биологически активными или биогенными аминами. Они являются медиаторами, с помощью которых сигнал передается от одной клетки к другой и от одной молекулы к другой. Субстратная специфичность декарбоксилаз очень разная.
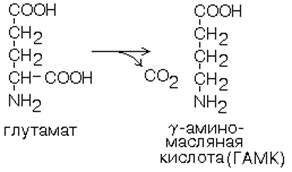
1. Глутаматдекарбоксилаза - высокоспецифичный фермент. Работает в клетках серого вещества головного мозга. Катализирует реакцию превращения глутаминовой кислоты в гамма-аминомасляную кислоту (ГАМК). ГАМК является медиатором тормозных импульсов в нервной системе. ГАМК и ее аналоги применяются в медицине как нейротропные средства для лечения эпилепсии и других заболеваний. 2. Орнитиндекарбоксилаза - высокоспецифичный фермент. Катализирует превращение орнитина в путресцин:
Образующийся путресцин (диаминобутан) является трупным ядом. В результате присоединения остатков пропиламина из путресцина могут образоваться спермин и спермидин, содержащие 3 (у спермина) или 4 (у спермидина) имино- или аминогруппы.
Полиамины принимают участие в процессах пролиферации клеток и роста тканей, а также в регуляции биосинтеза белка. Они являются ингибиторами некоторых ферментов, в том числе протеинкиназ. Спермин и спермидин относятся к группе биогенных полиаминов. Введение полиаминов в организм снижает температуру тела и кровяное давление.
При раковых заболеваниях обнаружено резкое увеличение секреции полиаминов и повышение их экскреции с мочой. Гистидиндекарбоксилаза Этот фермент имеет абсолютную субстратную специфичность - превращает гистидин в гистамин:
Гистамин является медиатором и содержится в нервных клетках и в тучных клетках. Обладают сильным сосудорасширяющим действием. Гистамин хранится в секреторных гранулах и секретируется в кровь при повреждении ткани (удар, ожог и т.д.). Особенно много его выделяется в очаге воспаления. Гистамин играет важную роль в проявлении аллергических реакций. Известно 2 типа рецепторов к гистамину:H1 и H2. Эффекты гистамина: - расширение капилляров и повышение сосудистой проницаемости; - понижение артериального давления; - повышение тонуса (спазм) гладких мышц - в том числе гладкой мускулатуры бронхов; - усиление секреции желудочного сока; Некоторые из этих эффектов позволяют гистамину принимать участие в формировании аллергических проявлений. Антигистаминные препараты применяются с целью предотвратить образование гистамина и обладают противовоспалительным и антиаллергическим действием. По механизму действия некоторые из них являются ингибиторами гистидиндекарбоксилазы, а другие конкурируют с гистамином за взаимодействие с рецепторами клеток. Блокаторы Н1-рецепторов используются в основном как противоаллергические средства - димедрол, супрастин и т.д. 4. Сериндекарбоксилаза. Катализирует первую реакцию синтеза ацетилхолина из серина. Ацетилхолин – возбуждающий медиатор вегетативной нервной системы. 5. Декарбоксилаза ароматических аминокислот Имеет широкую субстратную специфичность. Превращает несколько разных аминокислот: а) триптофан - в триптамин б) 5-окситриптофан - в триптамин (серотонин) в) 3,4-диоксифенилаланин - в дофамин
Серотонин вырабатывается в нервной ткани. Некоторые виды головных болей (мигрени) связаны с избыточной выработкой серотонина. Серотонин сужает сосуды, регулирует свертывание крови. Обладает антиаллергическим действием. Триптамин обладает сходным эффектом. Аминокислота фенилаланин может в результате окисления присоединять две ОН-группы в кольце и превращаться в диоксифенилаланин (ДОФА). Из него под действием декарбоксилазы ароматических аминокислот образуется дофамин. Дофамин является предшественником катехоламинов - норадреналина и адреналина.
Кроме функции предшественника, ДОФАмин имеет свои специфические функции.
ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ В организме имеются механизмы, позволяющие разрушать биогенные амины.
1. Метилирование по оксигруппам тех аминов, которые такие группы содержат, либо включают оксигруппы в свою молекулу после гидроксилирования. Источник метильного радикала: S-Аденозилметионин. Реакцию катализируют ферменты - О-метилтрансферазы. Они переносят метильную группу на кислород. После присоединения аденильного остатка АДФ к сере метионина, метильная группа метионина становится очень мобильной и легко переносится на разные вещества. В том числе и на кислород оксигрупп. 2. Окисление амина по аминогруппе с целью дезаминирования. Главный путь инактивации биогенных аминов - их окисление под действием оксидаз с отщеплением аминогруппы. В результате исчезает биологическая активность амина.
Оксидазы биогенных аминов: моноаминооксидаза (МАО), диаминооксидаза (ДАО), полиаминооксидаза. Оксидазы отнимают два протона и два электрона и передают их сразу на кислород. Образуется перекись водорода, а амин превращается в ИМИН. Этот имин легко гидролизуется без участия фермента и превращается в альдегид. Простетической группой ферментов оксидаз является ФАД или ФМН, т.е. они являются флавопротеинами. Вторая реакция (гидролиз) необратима. Образовавшийся в итоге альдегид легко окисляется до карбоновой кислоты, которая распадается до СО2 и H2O. МАО в клетке больше, чем ДАО. Угнетение МАО приволит к замедлению распада биогенных аминов. Такие лекарства продлевают период существования биогенных аминов, что особенно важно при их недостатке. Эти вещества играют роль антидепрессантов и используются, в частности, при лечении шизофрении. Аминокислоты декарбоксилируются в цитоплазме, а окисление аминов происходит в наружной мембране митохондрий. Поскольку реакция декарбоксилирования аминокислот и разрушение биогенных аминов происходят не одновременно, то биогенные амины могут некоторое время существовать и выполнять свою биологическую функцию. ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ У человека происходит в основном путем окислительного дезаминирования. Эти реакции протекают с помощью двух ферментов: - оксидаза Д-аминокислот - оксидаза L-аминокислот
Эти ферменты обладают групповой стереоспецифичностью. Оксидазы отнимают протоны и электроны от аминокислот с помощью такого же механизма, как и оксидазы, обеспечивающие дезаминирование биогенных аминов. Эти ферменты являются флавопротеинами и содержат в качестве простетической группы ФАД или ФМН: На первой стадии образуется иминокислота, а затем, после спонтанного гидролиза образуется альфа-кетокислота. Кроме оксидаз имеется еще один фермент, катализирующий окислительное дезаминирование глутаминовой кислоты - глутамат-дегидрогеназа (глутаматДГ). Этот фермент является НАД-зависимым и обладает высокой активностью (как и другие НАД-зависимые дегидрогеназы). В этом его отличие от оксидаз аминокислот, которые медленно превращают аминокислоты в физиологических условиях (поэтому в клетке сохраняется большинство аминокислот). Так как глутамат-ДГ является никотинамидной, то отнимаемые протоны и электроны не передаются сразу на кислород, а транспортируются по полной цепи МтО с образованием воды и параллельным образованием трех молекул АТФ.
Глутамат-ДГ обладает высокой активностью и этим отличается от МАО и ДАО. Глутамат-ДГ является регуляторным ферментом - он ингибируется избытком АТФ, и активируется избытком АДФ. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ 1. Реакции дезаминирования необратимы, как и реакции декарбоксилирования - дезаминирование тоже может играть роль первого этапа на путях распада аминокислот. 2. Один из непосредственных продуктов дезаминирования -конечный продукт метаболизма аммиак. Это токсическое вещество. Поэтому клетки должны затрачивать энергию, чтобы обезвредить аммиак до безвредных продуктов, которые выводятся из организма. 3. Другой продукт реакции дезаминирования - альфа-кетокислота. Все образующиеся альфа-кетокислоты легко расщепляются дальше до СО2 и Н2О (например, аланин превращается в ПВК (путем дезаминирования; аспартат - в ЩУК; глутаминовая кислота - в альфа-кетоглутаровую). Большинство альфа-кетокислот тем или иным путем превращаются в кислоты, которые являются промежуточными метаболитами ЦТК: - в альфа-кетоглутаровую; - в янтарную; - фумаровую; -щавелево-уксусную. Все эти метаболиты могут в организме трансформироваться в углеводы, перед этим превращаясь в ПВК. Поэтому большинство аминокислот относится к группе, которая называется глюкогенными аминокислотами (их 17). Только 3 аминокислоты не могут превращаться в пируват, но превращаются в Ацетил-КоА – кетогенные аминокислоты: лейцин, лизин, триптофан). Они могут прямо трансформироваться в жирные кислоты или в кетоновые тела.
Метаболические пути, в которые вступают аминокслоты после дезаминирования, уже не являются собственно путями метаболизма аминокислот, а являются универсальными и для аминокислот, и для углеводов, и для жиров. ТРАНСАМИНИРОВАНИЕ Эта реакция заключается в том, что аминокислота и кетокислота обмениваются друг с другом своими функциональными группами при альфа-углеродном атоме. В результате вступившая в реакцию аминокислота превращается в соответствующую альфа-кетокислоту, а кетокислота становится аминокислотой. Эту реакцию катализируют ферменты под названием ТРАНСАМИНАЗЫ (АМИНОТРАНСФЕРАЗЫ). Коферментом всех трансаминаз является активная форма витамина В6 - пиридоксальфосфат (фосфопиридоксаль). ГЛАВНЫЕ ОСОБЕННОСТИ РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ: 1. Это циклический процесс, все стадии которого катализируются одним и тем же ферментом - трансаминазой данной пары кислот. В этот цикл вступает одна аминокислота и кетокислота. Образуются другая альфа-кетокислота и аминокислота. 2. Все стадии этого процесса обратимы. Поэтому весь цикл в целом может протекать как в прямом, так и в обратном направлении. Направление этого цикла зависит от соотношения концентраций участников реакции - всех четырех кислот. Резко повысив концентрацию какой-либо одной из кислот, можно направить реакцию по определенному направлению. 3. Каждая трансаминаза обычно специфична для одной пары субстратов и соответствующей ей пары продуктов и все стадии реакции катализируются только одним ферментом. По участникам процесса для данного фермента дают ему название:
Фермент, который катализирует приведенную реакцию, можно назвать: Аланинаминотрансфераза (АлТ) (глутаминово-пировиноградная трансаминаза (ГПТ)). 4. Трансаминазы обычно обладают высокой субстратной специфичностью и высокой активностью. Наиболее активными в клетках являются те трансаминазы, для которых хотя бы один из субстратов является дикарбоновой альфа-кетокислотой - это АлТ - аланинаминотрансфераза (ГПТ) и АсТ - аспартатаминотрансфераза (ГЩТ). Определение активности этих ферментов в сыворотке крови имеет важное значение в ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКЕ таких заболеваний, как ВИРУСНЫЙ ГЕПАТИТ и ИНФАРКТ МИОКАРДА. АсТ и АлТ являются внутриклеточными ферментами. Поэтому в норме их активность в крови очень мала. При вирусном гепатите или при инфаркте миокарда наблюдается разрушение клеток печени или миокарда соответственно. Поэтому в крови будет наблюдаться повышение активности и АлТ, и АсТ, но неодинаковое для каждого из ферментов. В клетках сердечной мышцы (миокарда) содержится гораздо больше АсТ, чем АлТ, а в клетках печени - наоборот: АлТ намного больше, чем АсТ. Поэтому ПРИ ИНФАРКТЕ МИОКАРДА АКТИВНОСТЬ АСТ в крови БУДЕТ ЗНАЧИТЕЛЬНО ВЫШЕ, ЧЕМ АЛТ, А ПРИ ВИРУСНОМ ГЕПАТИТЕ АКТИВНОСТЬ АЛТ БУДЕТ ВЫШЕ, ЧЕМ АСТ. Поэтому определение активности этих ферментов в крови помогает врачам в постановке правильного диагноза. БИОЛОГИЧЕСКАЯ РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ. 1. ОБЕСПЕЧИВАЮТ СИНТЕЗ НОВЫХ АМИНОКИСЛОТ ИЗ ЧИСЛА ЗАМЕНИМЫХ. Из заменимых аминокислот также могут образоваться необходимые клетке кетокислоты. Эта функция позволяет ругулировать содержание различных аминокислот в клетках организма (корректировка аминокислотного состава клеток). При избытке какой-либо из кислот соотношение быстро восстанавливается трансаминазой за счет другой пары. Недостающее количество какой-либо из кислот может быть взято клеткой из других метаболических путей (например, альфа-кетоглутарат может быть взят из ЦТК). Избыток какой-либо из кислот может быть ликвидирован другими ферментами (например, избыточное количество глутамата окисляется глутаматдегидрогеназой). 2. ОБЕСПЕЧИВАЮТ ПРОТЕКАНИЕ РЕАКЦИЙ КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ. 3. ОБЕСПЕЧИВАЮТ СИНТЕЗ МОЧЕВИНЫ
ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ Катаболизм аминокислот ускоряется при: поступлении с пищей больших количеств белка, гак как аминокислоты не могут запасаться в организме; голодании, так как распадаются белки тканей; сахарном диабете и других длительно протекающих тяжелых заболеваниях. Катаболизм аминокислот чаще всего начинается с реакции дезаминирования - удаления a-аминогруппы. В отличие от трансаминирования при дезаминировании общее количество аминокислот уменьшается. a -Аминогруппа отщепляется в виде аммиака. Аммиак - токсичное соединение, поэтому в клетках подвергается обезвреживанию. Безазотистый остаток представляет собой a --кетокислоту, которая включается (рис.3): • в реакции трансаминирования для синтеза заменимых аминокислот; • в анаплеротические реакции для восполнения убыли метаболитов ОПК, используемых для синтеза других соединений; • в глюконеогенез; • в кетогенез; • в реакции окисления до СО2, и Н20.
Рисунок 3 Дезаминированию подвергаются все аминокислоты, кроме лизина. Реакции дезаминирования: а) окислительное - для Глу; б) неокислительное - для Сер, Тре, Гис и Цис; в) непрямое - для остальных аминокислот. Окислительное дезаминирование глутамата происходит в присутствии кофермента NAD+. Реакция происходит в митохондриях многих тканей, наиболее активно в печени. Прямому окислительному дев минированию подвергается только глутамат. Фермент, катализирующий эту реакцию глутаматдегидрогеназа. Г лутамат-ДГ является никотинамидной, то отнимаемые протоны и электроны не передаются сразу на кислород, а транспортируются по полной цепи МтО с образованием воды и параллельным образованием трех молекул АТФ. Прямому неокислительному дезаминированию подвергаются: • серии и треонин - дезаминирование происходит с отщеплением воды; • гистидин дезаминируется внутримолекулярным способом; • цистеин дезаминируется с выделением H2S и с использованием Н2О. Большинство аминокислот подвергается в клетке непрямому дезаминированию, которое включает двестадии: • трансаминирование с a - -кетоглутаратом, образование Глу в цитозоле клетки; • окислительное дезаминирование Глу в митохондриях. Центральную роль в непрямом дезаминированини иг рают глутамат и a -кетоглутарат. ОБМЕН АММИАКА Основным источником аммиака является катаболизм аминокислот, однако аммиак может образовываться при распаде других азотсодержащих соединений(биогенных аминов, нуклеотидов) в тканях (рис.4). Часть аммиака образуется в кишечнике в результате жизнедеятельности микрофлоры кишечника (гниение белков) и всасывается в кровь воротной вены. Концентрация аммиака в крови воротной вены существенно выше, чем в общем кровотоке.
Рисунок 4 Катаболизм аминокислот и, следовательно, образование аммиака происходят во всех тканях. Концентрация аммиака в крови очень мала, так как он является токсичным веществом и быстро обезвреживается в клетках. Содержание аммиака в крови в норме 0.4 -0,7мг/л, или 25-40 мкмоль/л. Токсичность аммиака связана с его действием на центральную нервную систему. Образовавшийся в клетках аммиак связывается (обезвреживается) и выводится почками в виде конечных продуктов азотистого обмена: • мочевины - синтезируется в печени; • аммонийных солей - образуются в почках. Существует несколько способов обезвреживания и выведения аммиака в разных тканях (рис.5). Основной реакцией обезвреживания аммиака почти во всех тканях является синтез глутамина действием глутаминсинтетазы: Глутаминсинтетаза обладает высоким сродством к аммиаку, и благодаря этой реакции в крови и тканях поддерживается низкая концентрация NH3. Глутамин является транспортной формой аммиака, так как представляет собой нейтральную аминокислоту, способную легко проникать через клеточные мембраны путем облегченной диффузии (в отличие от глутаминовой кислоты, требующей механизмов активного транспорта). Из многих органов глутамин поступает в кровь, в наибольшем количестве - из мышц и мозга. Образовавшийся в тканях глутамин транспорти руется в почки и кишечник. В клетках кишечника Глн отщепляется амидная группа в виде NH3, образовавшийся глутамат трансаминируется с образованием аланина. В энтероцитах амидная группа глутамина превращается в аммиак, а аминогруппа глутамина - в аминогруппу аланина. В почках глутамин также подвергается действию глутаминазы и расщепляется на аммиак и глутамат, который реабсорбируется и аозвращается в клетки тканей. Глутаминаза почек активируется при ацидозе, когда полученный аммиак используется для нейтрализации кислых продуктов и образования аммонийных солей, которые экскретируются с мочой. Эта реакция защищает организм от потери с мочой ионов натрия и калия, которые также могут быть использованы для выведения избытка анионов. Еще одной реакцией обезвреживания аммиака в мозге и некоторых других органах является восстановительное аминирование a-кетоглутарата под действием глутаматдегидрогеназы, катализирующей обратимую реакцию. Однако этот путь обезвреживания аммиака в тканях используется слабо, хотя, если учитывать последующее образование глутамина, является выгодным для клеток, так как способствует обезвреживанию сразу 2 молекул NH3 Из мышц и некоторых других тканей избыток аммиака выводится в виде аланина. Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, источником которого служат безазотистые остатки аминокислот, а также глюкоза. Особенно много аланина выделяют мышцы в силу их большой массы, а также потому, что работающие мышцы часть энергии получают за счет распада аминокислот
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 495; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.36.215 (0.014 с.) |