Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Внешнее и внутреннее строение лишайниковСодержание книги Поиск на нашем сайте
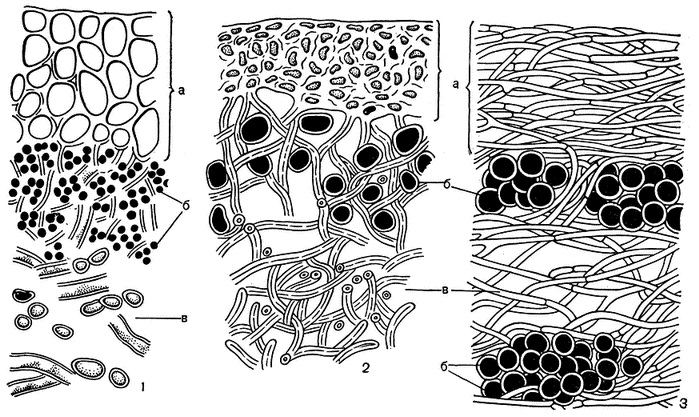
Вегетативное тело лишайника — таллом, или слоевище, очень разнообразно по форме и окраске. Лишайники окрашены в самые различные цвета: белый, розовый, ярко-желтый, оранжевый, оранжево-красный, серый, голубовато-серый, серовато-зеленый, желтовато-зеленый, оливково-коричневый, коричневый, черный и некоторые другие. Окраска слоевища лишайников зависит от наличия пигментов, которые откладываются в оболочках гиф, реже в протоплазме. Наиболее богаты пигментами гифы корового слоя лишайников и различные части их плодовых тел. У лишайников различают пять групп пигментов: зеленые, синие, фиолетовые, красные, коричневые. Механизм образования их до сих пор не выяснен, но совершенно очевидно, что важнейшим фактором, влияющим на этот процесс, является свет. Иногда цвет слоевища зависит от окраски лишайниковых кислот, которые откладываются в виде кристаллов или зернышек на поверхности гиф. Большинство лишайниковых кислот бесцветны, но некоторые из них окрашены, и иногда очень ярко – в желтый, оранжевый, красный и другие цвета. Окраска кристаллов этих веществ определяет и окраску всего слоевища. И здесь важнейшим фактором, способствующим образованию лишайниковых веществ, является свет. Чем ярче освещение в месте произрастания лишайника, тем ярче он окрашен. Как правило, очень ярко окрашены лишайники высокогорий и полярных районов Арктики и Антарктики. Это тоже связано с условиями освещения. Для высокогорных и полярных районов земного шара характерны большая прозрачность атмосферы и высокая интенсивность прямой солнечной радиации, обеспечивающие здесь значительную яркость освещения. В таких условиях в наружных слоях слоевищ концентрируется большое количество пигментов и лишайниковых кислот, обусловливая яркую окраску лишайников. Предполагают, что окрашенные наружные слои защищают нижележащие клетки водорослей от чрезмерной интенсивности освещения. Из-за низкой температуры осадки выпадают в Антарктике только в виде снега. В такой форме они не могут быть использованы растениями. Вот здесь-то темная окраска лишайников и приходит им на помощь. Темноокрашенные слоевища антарктических лишайников за счет высокой солнечной радиации быстро нагреваются до положительной температуры даже при отрицательной температуре воздуха. Снег, падающий на эти нагретые слоевища, тает, превращаясь в воду, которую лишайник сразу же впитывает. Таким образом он обеспечивает себя водой, необходимой для осуществления процессов дыхания и фотосинтеза. Насколько разнообразны слоевища лишайников по окраске, настолько же разнообразны они и по форме. Слоевище может иметь вид корочки, листовидной пластинки или кустика. В зависимости от внешнего облика различают три основных морфологических типа: На́кипные. Таллом накипных лишайников — это корочка («накипь»), нижняя поверхность плотно срастается с субстратом и не отделяется без значительных повреждений. Это позволяет им жить на крутых склонах гор, деревьях и даже на бетонных стенах. Иногда накипный лишайник развивается внутри субстрата и снаружи совершенно не заметен. Как правило, накипные слоевища небольших размеров, их диаметр составляет всего несколько миллиметров или сантиметров, но иногда может достигать и 20 – 30 см. В природе нередко можно наблюдать, как небольшие по размерам накипные слоевища лишайников, сливаясь друг с другом, образуют на каменистой поверхности скал или стволах деревьев крупные пятна, достигающие в диаметре нескольких десятков сантиметров. Листоватые. Листоватые лишайники имеют вид пластин разной формы и размера. Они более или менее плотно прикрепляются к субстрату при помощи выростов нижнего коркового слоя. Наиболее простое слоевище листоватых лишайников имеет вид одной крупной округлой листовидной пластинки, достигающей в диаметре 10 – 20 см. Такая пластинка нередко бывает плотной, кожистой, окрашенной в темно серый, темно-коричневый или черный цвет. Кустистые. По организационному уровню кустистые лишайники представляют высший этап развития слоевища. У кустистых лишайников таллом образует множество округлых или плоских веточек. Растут на земле или свисают с деревьев, древесных остатков, скал. Слоевище кустистых лишайников имеет вид прямостоячего или повисающего кустика, реже неразветвленных прямостоячих выростов. Это позволяет кустистым лишайникам путем изгибов веточек в разные стороны занимать наилучшее положение, при котором водоросли могут максимально использовать свет для осуществления фотосинтеза. Слоевища кустистых лишайников могут быть разных размеров. Высота самых маленьких составляет всего несколько миллиметров, а наиболее крупных 30 – 50 см. Повисающие слоевища кустистых лишайников иногда могут достигать колоссальных размеров. Внутреннее строение лишайника: коровый слой, гонидиальный слой, сердцевина, нижняя кора, ризоиды. Тело лишайников (таллом) представляет собой переплетение грибных гиф, между которыми находится популяция фотобионта.
1 - гетеромерное слоевище (а - верхний коровой слой, б - слой водорослей, в - сердцевина, г - нижний коровой слой); 2 - гомеомерное слоевище слизистого лишайника коллема (Collema flaccidum); 3 - гомеомерное слоевище слизистого лишайника лептогиум (Leptogium saturninum) (а - коровой слой с верхней и нижней стороны слоевища, б - ризоиды) Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника определенную функцию и в зависимости от этого имеет совершенно определенное строение. Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид очень тонких нитей, состоящих из одного ряда клеток. Эти нити называют ризоидами. Каждая такая нить берет начало от одной клетки нижнего корового слоя. Нередко несколько ризоидов соединяются в толстые ризоидальные тяжи. В зоне водорослей осуществляются процессы ассимиляции углекислоты и накопление органических веществ. Как известно, для осуществления процессов фотосинтеза водорослям не обходим солнечный свет. Поэтому слой водорослей обычно размещается вблизи верхней поверхности слоевища, непосредственно под верхним коровым слоем, а у вертикально стоящих кустистых лишайников еще и над нижним коровым слоем. Слой водорослей чаще всего бывает небольшой толщины, и водоросли размещаются в нем так, что находятся почти в одинаковых условиях освещения. Водоросли в слоевище лишайника могут образовывать непрерывный слой, но иногда гифы микобионта делят его на отдельные участки. Для осуществления процессов ассимиляции углекислоты и дыхания водорослям необходим также нормальный газообмен. Поэтому грибные гифы в зоне водорослей не образуют плотных сплетений, а расположены рыхло на некотором расстоянии друг от друга. Под слоем водорослей расположен сердцевинный слой. Обычно сердцевина по толщине значительно превышает коровой слой и зону водорослей. От степени развития сердцевины зависит толщина самого слоевища. Основная функция сердцевинного слоя – проведение воздуха к клеткам водорослей, содержащим хлорофилл. Поэтому для большинства лишайников характерно рыхлое расположение гиф в сердцевине. Воздух, попадающий в слоевище, легко проникает к водорослям по промежуткам между гифами. Сердцевинные гифы слабоветвисты, с редкими поперечными перегородками, с гладкими, слабожелатинообразными толстыми стенками и довольно узким просветом, заполненным протоплазмой. У большинства лишайников сердцевина белая, так как гифы сердцевинного слоя бесцветны. По внутреннему строению лишайники разделяют на: — гомеомерные (Collema), клетки фотобионта распределены хаотично среди гиф гриба по всей толщине таллома; — гетеромерные (Peltigera canina), таллом на поперечном срезе можно чётко разделить на слои. Лишайников с гетеромерным талломом большинство. В гетеромерном талломе верхний слой — корковый, сложенный гифами гриба. Он защищает таллом от высыхания и механических воздействий. Следующий от поверхности слой — гонидиальный, или альгальный, в нём располагается фотобионт. В центре располагается сердцевина, состоящая из беспорядочно переплетенных гиф гриба. В сердцевине в основном запасается влага, она также играет роль скелета. У нижней поверхности таллома часто находится нижняя кора, с помощью выростов которой (ризин) лишайник прикрепляется к субстрату. Полный набор слоёв встречается не у всех лишайников. Как и в случае двухкомпонентных лишайников, водорослевый компонент — фикобионт — трёхкомпонентных лишайников равномерно распределен по таллому, либо образует слой под верхней корой. Некоторые трёхкомпонентные цианолишайники образуют специализированные поверхностные или внутренние компактные структуры (цефалодии), в которых сосредоточен цианобактериальный компонент. Способы питания лишайников Лишайники представляют для физиологических исследований сложный объект, так как состоят из двух физиологически противоположных компонентов – гетеротрофного гриба и автотрофной водоросли. Поэтому приходится сначала отдельно изучать жизнедеятельность мико- и фикобионта, что делается с помощью культур, а затем жизнь лишайника как целостного организма. Понятно, что такая «тройная физиология» – трудный путь исследования, и не удивительно, что в жизнедеятельности лишайников кроется еще много загадочного. Однако общие закономерности их обмена веществ все же выяснены. Довольно много исследований посвящено процессу фотосинтеза у лишайников. Так как лишь небольшая часть их слоевища (5 – 10% объема) образована водорослью, которая тем не менее является единственным источником снабжения органическими веществами, встает существенный вопрос об интенсивности фотосинтеза в лишайниках. Как показали измерения, интенсивность фотосинтеза у лишайников намного ниже, чем у высших автотрофных растений. Для нормальной фотосинтетической активности слоевище должно содержать определенное количество воды, зависящее от анатомоморфологического типа лишайника. В общем в толстых слоевищах оптимальное содержание воды для активного фотосинтеза ниже, чем в тонких и рыхлых слоевищах. При этом весьма существенно то обстоятельство, что многие виды лишайников, особенно в сухих местообитаниях, вообще редко или по крайней мере очень нерегулярно снабжаются оптимальным количеством внутрислоевищной воды. Ведь регуляция водного режима у лишайников происходит совсем по-иному, чем у высших растений, имеющих специальный аппарат, способный контролировать получение и расходование воды. Лишайники усваивают воду (в виде дождя, снега, тумана, росы и пр.) очень быстро, но пассивно всей поверхностью своего тела и отчасти ризоидами нижней стороны. Такое поглощение слоевищем воды представляет собой простой физический процесс, как, например, впитывание воды фильтровальной бумагой. Лишайники способны впитывать воду в очень больших количествах, обычно до 100 – 300% от сухой массы слоевища, а некоторые слизистые лишайники (коллемы, лептогиумы и др.) даже до 800 – 3900%. Минимальное содержание воды в лишайниках в природных условиях составляет примерно 2 – 15% от сухой массы слоевища. Отдача воды слоевищем также происходит довольно быстро. Насыщенные водой лишайники на солнце через 30 – 60 мин теряют всю свою воду и делаются хрупкими, т. е. содержание воды в слоевище становится ниже минимально необходимого для активного фотосинтеза. Из этого вытекает своеобразная «аритмичность» фотосинтеза лишайников – его продуктивность меняется в течение дня, времени года, ряда годов в зависимости от общих экологических условий, особенно гидрологических и температурных. Имеются наблюдения, что многие лишайники более активно фотосинтезируют в утренние и вечерние часы и что фотосинтез продолжается у них и зимой, а у напочвенных форм даже под нетолстым снеговым покровом. Важным компонентом в питании лишайников является азот. Те лишайники, которые имеют в качестве фикобионта зеленые водоросли (а их большинство), воспринимают соединения азота из водных растворов, когда их слоевища пропитываются водой. Возможно, что часть азотистых соединений лишайники берут и прямо из субстрата – почвы, коры деревьев и т. д. Экологически интересную группу составляют так называемые нитрофильные лишайники, растущие в местообитаниях, богатых азотистыми соединениями, – на «птичьих камнях», где много экскрементов птиц, на стволах деревьев и т. д. (виды ксантории, фисции, калоплаки и др.). Лишайники, имеющие в качестве фикобионта сине-зеленые водоросли (особенно ностоки), способны фиксировать атмосферный азот, так как этой способностью обладают содержащиеся в них водоросли. В опытах с такими видами (из родов коллема, лептогиум, пельтигера, лобария, стикта и др.) было установлено, что их слоевища быстро и активно поглощают атмосферный азот. Эти лишайники часто селятся на субстратах, весьма бедных азотистыми соединениями. Большая часть азота, фиксированного водорослью, направляется микобионту и лишь незначительная часть используется самим фикобионтом. Имеются данные, что микобионт в слоевище лишайника ведет активный контроль над освоением и распределением азотистых соединений, фиксированных из атмосферы фикобионтом. Описанный выше ритм жизни является одной из причин для очень медленного роста большинства лишайников. Иногда лишайники растут всего лишь на несколько десятых миллиметра в год, в основном менее чем на один сантиметр. Другой причиной медленного роста является то, что фотобионт, составляя нередко менее 10% объёма лишайника, берёт на себя обеспечение микобионта питательными веществами. В хороших условиях, с оптимальными влажностью и температурой, например в туманных или дождливых тропических лесах, лишайники растут на несколько сантиметров в год. Ростовая зона лишайников у накипных форм находится по краю лишайника, у листоватых и кустистых на каждой верхушке. Лишайники являются одними из самых долгоживущих организмов и могут достигать возраста нескольких сотен лет, а в некоторых случаях более 4500 лет, как например Rhizocaгрon geographicum, живущий в Гренландии. Размножение лишайников Лишайники размножаются либо спорами, которые образует микобионт половым или бесполым путем, либо вегетативно – фрагментами слоевища, соредиями и изидиями. При половом размножении на слоевищах лишайников в результате полового процесса формируются половые спороношения в виде плодовых тел. Среди плодовых тел у лишайников различают апотеции, перитеции и гастеротеции. Большинство лишайников формируют открытые плодовые тела в виде апотециев – дисковидных образований. Некоторые имеют плодовые тела в форме перитеция – закрытого плодового тела, имеющего вид маленького кувшина с отверстием наверху. Небольшое количество лишайников образуют узкие плодовые тела удлиненной формы, которые называют гастеротециями. В апотециях, перитециях и гастеротециях споры развиваются внутри сумок – особых мешковидных образований. Лишайники, формирующие споры в сумках, объединяются в большую группу сумчатых лишайников. Они произошли от грибов класса аскомицетов и представляют основную эволюционную линию развития лишайников. У небольшой группы лишайников споры образуются не внутри сумок, а экзогенно, на вершине удлиненно-булавовидных гиф – базидий, на концах которых развиваются четыре споры. Лишайники с таким образованием спор объединяются в группу базидиальных лишайников. Женский половой орган лишайников – архикарп – состоит из двух частей. Нижняя часть носит название аскогона и представляет собой спирально закрученную гифу, более толстую по сравнению с другими гифами и состоящую из 10 – 12 одно- или много ядерных клеток. От аскогона вверх отходит трихогина – тоненькая вытянутая гифа, которая проходит через зону водорослей и коровой слой и выходит на поверхность слоевища, возвышаясь над ней своей липкой верхушкой. Развитие и созревание плодового тела у лишайников – очень медленный процесс, который длится 4 – 10 лет. Сформировавшееся плодовое тело тоже является многолетним, способным в течение ряда лет продуцировать споры. Сколько же спор способны продуцировать плодовые тела лишайников? Подсчитано, например, что у лишайника солорина в апотеции диаметром 5 мм образуется 31 тыс. сумок, а в каждой сумке обычно развивается по 4 споры. Следовательно, общее количество спор, продуцируемое одним апотецием, равно 124 000. В течение одного дня из такого апотеция выбрасывается от 1200 до 1700 спор. Конечно, не все выброшенные из плодового тела споры прорастают. Многие из них, попав в неблагоприятные условия, погибают. Для прорастания споры необходимы прежде всего достаточная влажность и определенная температура. У лишайников известны также бесполые спороношения – конидии, пикноконидии и стилоспоры, возникающие экзогенно на поверхности конидиеносцев. При этом конидии образуются на конидиеносцах, развивающихся непосредственно на поверхности слоевища, а пикноконидии и стилоспоры в особых вместилищах – пикнидиях. Из бесполых спороношений лишайники чаще всего формируют пикнидии с пикноконидиями. Пикнидии нередко встречаются на слоевищах многих кустистых и листоватых лишайников, реже их можно наблюдать у накипных форм. В каждом из пикнидиев образуются в огромном количестве маленькие одноклеточные споры – пикноконидии. Роль этих столь широко распространенных спороношений в жизни лишайника до сих пор не выяснена. Одни ученые, называя эти споры спермациями, а пикнидии – спермагониями, считают их мужскими половыми клетками, хотя до сих пор нет ни экспериментальных, ни цитологических данных, доказывающих, что пикноконидии действительно участвуют в половом процессе лишайников. Вегетативное размножение. Если накипные лишайники, как правило, образуют плодовые тела, то среди более высокоорганизованных листоватых и кустистых лишайников имеется немало представителей, которые размножаются исключительно вегетативным путем. В этом случае более важны для размножения лишайников такие образования, в которых одновременно присутствуют гифы гриба и клетки водоросли. Это соредии и изидии. Они служат для размножения лишайника как целого организма. Попав в благоприятные условия, они дают начало непосредственно новому таллому. Соредии и изидии встречаются чаще у листоватых и кустистых лишайников. Соредии представляют собой мельчайшие образования в виде пылинок, состоящих из одной или нескольких клеток водоросли, окруженных гифами гриба. Формирование их обычно начинается в гонидиальном слое. Вследствие массового образования соредий количество их увеличивается, они давят на верхнюю кору, разрывают ее и оказываются на поверхности таллома, откуда легко сдуваются при любом движении воздуха или смываются водой. Скопления соредий называют соралями. Наличие и отсутствие соредий и соралей, их расположение, форма и окраска постоянны для определенных лишайников и служат определительным признаком. Иногда при отмирании лишайников их таллом превращается в порошковатую массу, состоящую из соредий. Это так называемые лепрозные формы лишайников (от греческого слова «лепрос» — «шероховатый», «неровный»). В этом случае определить лишайник почти не представляется возможным. Соредии, разносимые ветром и дождевой водой, попав в благоприятные условия, постепенно образуют новый таллом. Возобновление нового таллома из соредии происходит очень медленно. Так, у видов из рода кладония нормальные чешуйки первичного таллома развиваются из соредии только через срок от 9 до 24 месяцев. А для развития вторичного таллома с апотециями требуется от одного до восьми лет в зависимости от вида лишайника и внешних условий. Изидии встречаются у меньшего числа видов лишайников, нежели соредии и сорали. Они представляют собой простые или коралловидно разветвленные выросты, обычно густо покрывающие верхнюю сторону таллома (см. рисунок). В отличие от соралей изидии снаружи покрыты корой, часто более темной, чем таллом. Внутри, под корой, они содержат водоросли и грибные гифы. Изидии легко отламываются от поверхности таллома. Обламываясь и распространяясь с помощью дождя и ветра, они, так же как и соредии, могут при благоприятных условиях образовывать новые талломы лишайников. Многие лишайники не образуют апотециев, соредии и изидии и размножаются участками таллома, которые легко отламываются от хрупких в сухую погоду лишайников ветром или животными и ими же переносятся. Особенно широко распространено размножение лишайников участками таллома в арктических областях, представители родов цетрария и кладония, многие из которых почти никогда не образуют плодовых тел. Значение: корм для животных, почвообразовательный процесс, для получения красителей Это своеобразные симбиотические организмы, состоящие из гриба и водоросли. Известно около 20000 лишайников. Их двойственная природа была установлена в середине 19 века немецким ученым С. Шведенером. Несмотря на разнообразие внешней формы, все они имеют сходное внутреннее строение: тело лишайника, слоевище, образовано гифами гриба, между которами распологаются зеленые или синезеленые водоросли. В этом симбиозе гриб защищает водоросли от высыхания, механического воздействия и действия очень высоких и очень низких температур. Он обеспечивает водоросль раствором минеральных солей. В свою очередь от водоросли получает синтезируемые ею углеводы, витамины группы В. Этот симбиоз не является гармоническим сосуществованием, а здесь происходит умеренный паразитизм гриба на водоросли. Лишайниковое сожительство постоянно и исторически обусловлено, а не случайно, кратковременно. Гриб способен образовывать присоски, которыми внедряется в клетки водоросли и поглощает их содержимое. В результате этого образуется организм, отличающийся новыми биологическими свойствами. Это обеспечивает существование лишайника в самых неблагоприятных условиях, где отдельно ни тот, ни другой организм не смогли бы развиваться (на голых скалах, бесплодных почвах, на стенах). Они широко распространены в тундре - арктической и высокогорной, в тайге - в сосновых борах, часто серым ковром выстилают почву, растут на стволах деревьев, свисают с ветвей (бородачи). Встречаются на лугах, в степях, в пустынях. Лишайники не имеют типичной зеленой окраски, у них нет стебля, листьев (этим они отличаются от мхов). Тело их состоит из слоевища, или таллома. Цвет лишайников зависит от окраски лишайниковых кислот, от пигментов, находящихся в оболочках гиф гриба
Лишайники размножаются вегетативным, бесполым и половым путём. Питаются при помощи фотосинтеза.
40. Отдел хвощевидные. Особенности строения и цикла развития на примере хвоща полевого. Роль в природе и жизни человека. Хвощи произошли от проптеридофитов. Хвощи – многолетние травы лугов, болот, лесов и полей. От корневищ отходят придаточные корни и надземные побеги. Побеги имеют членистое строение. От узлов отходят мутовки бурых чешуевидных листьев, сросшихся в трубчатое влагалище, и мутовки боковых побегов. Функцию фотосинтеза выполняют ребристые стебли. Клетки эпидермы пропитаны кремнеземом. Под эпидермой расположена механическая ткань. Коллатеральные проводящие пучки образуют кольцо. В ксилеме – различного типа трахеиды, иногда сосуды; флоэма состоит из ситовидных элементов и паренхимы. Место сердцевины в междоузлиях занято широкой полостью. Споры образуются в спороносных колосках, развивающихся на ассимилирующих или на специализированных спороносных бесхлорофилльных побегах. На оси колоска находятся спорангиофоры (щитки). После созревания спор щитки подсыхают и раздвигаются, стенки спорангия разрушают и споры освобождаются. Образовавшиеся в результате мейоза гаплоидные споры одеты толстой оболочкой. При прорастании образуются мужские и обоеполые заростки. У хвоща полевого весенние бесхлорофилльные неветвящиеся спороносные побеги, на верхушках которых находятся спороносные колоски, после созревания спор отмирают, а на смену им от тех же корневищ образуются летние вегетативные ассимилирующие побеги с мутовками зеленых веточек. Хвощ полевой – сорняк. При попадании в большом количестве в сено могут вызвать отравление животных. В медицине используют в качестве мочегонного средства при отеках. 41. Спороносные листочки (спорофиллы) плауновидных по форме, размерам и цвету могут быть похожи на вегетативные листья (трофофиллы) или в той или иной степени отличаться от них. Чередуясь со стерильными листьями, спорофиллы образуют на протяжении стебля спороносные зоны или собраны в расположенные на концах ветвей компактные образования – стробилы. У некоторых ископаемых форм плауновидных спорофиллы сидели на стебле вперемежку с вегетативными листьями, не образуя ни спороносных зон, ни стробилов. Гаметофит плаунов - многолетнее бесхлорофильное растение в виде клубенька с ризоидами. Заросток обоеполый, в глубине его ткани образуются архегонии и антеридии. Оплодотворение происходит при наличии воды. После оплодотворения из яйцеклетки развивается зародыш, вырастающий в многолетнее вечнозеленое растение - спорофит При этом они развиваются в подземных или полуподземных условиях, и питаются в значительной степени или исключительно за счет образования союза с почвенными грибами. Заростки разноспоровых плауновидных (к ним относятся все представители класса полушниковых) развиваются обычно в течение нескольких недель. Они вообще лишены хлорофилла и живут за счет питательных веществ, содержащихся в споре. Поэтому они очень мелкие и даже в зрелом виде лишь слегка выступают за пределы оболочки споры. В настоящее время на нашей планете встречаются представители двух классов отдела плауновидных – плауновых и полушниковых, или шильниковых. Все виды плаунов находятся под охраной как древние, вымирающие растения. 42. Голосемянные растения, у которых не бывает плодов и семена лежат открыто, голо. К ним относятся хвойные растения, которые имеют хвою (это видоизмененные листья) Строение: Корневая система стержневая (сосна) или мочковатая (ель) Стебель с тонкой корой и мощной древесиной. Листья видоизмененные в иглы или чашуйки, они расположены на уделенных или укороченных побегах, листья многолетнии (искл.лиственница) Размножение: половое – семенами. Размножение голосеменных. Сосна, как и все хвойные растения, размножается семенами. На чешуях раскрывшихся шишек семена расположены по два и лежат открыто, поэтому относят к голосеменным растениям. Размножение семенами — основной признак, по которому голосеменные отличаются от растений, размножающихся спорами. Голосеменные растения плода не образуют. Весной на ее молодых ветвях видны маленькие шишки. Одни из них зеленовато-желтые. Они собраны тесными группами у оснований молодых побегов. Другие красноватые, одиночные. Каждая шишка состоит из оси и сидящих на ней чешуй. На чешуях зеленоватых шишек развиваются по два пыльцевых мешочка. В них созревает пыльца. Оболочка каждой пылинки имеет два пузырька, наполненных воздухом. Такие пылинки ветер переносит на большие расстояния. · Красноватые шишки сосны находятся они на вершинах молодых веток. На чешуях красноватых шишек развиваются семязачатки (семяпочки), по два на чешуе. Созревшая пыльца высыпается, и ее подхватывает ветер. Опыление произведут те пылинки, которые будут перенесены на пыльцевходы семязачатков. После опыления семязачатков чешуи красноватых шишек смыкаются и склеиваются смолой. В пылинках образуются мужские гаметы, а в семязачатках — женские. Оплодотворение происходит в семязачатках закрывшихся шишек. Из зиготы развивается зародыш, из всего семязачатка — семя, а шишки растут и древеснеют. Сначала они зеленые, затем коричневые. · Семена у сосны созревают через полтора года после опыления, а высыпаются из шишек почти через два года. В семенах голосеменных растений имеется ткань, содержащая запас питательных веществ. Эта ткань окружает зародыш. Семена сосны имеют пленчатые крылышки, благодаря которым они распространяются ветром. Но у семян некоторых сосен крылышек нет. Семена сосны сибирской («кедровой сосны») называют «кедровыми орешками». Шишки сосны обыкновенной небольшие, длиной всего 4—5 см. Еловые шишки крупнее, длиной 10—15 см. Семена в шишках ели созревают осенью того же года, когда произошло опыление. В январе — марте созревшие семена высыпаются, подхватываются ветром и скользят по ледяному насту. Значение голосеменных. Ель, сосна, пихта, лиственница, можжевельник и все другие голосеменные растения имеют большое значение в природе и в жизни человека. Вместе с другими зелеными растениями они образуют органические вещества, усваивают углекислый газ из воздуха и выделяют кислород. Естественное заселение лесных вырубок и пожарищ, заброшенных полей начинается с появления на этих участках светолюбивой, неприхотливой сосны, березы, осины. Поселившись на сыпучих песках, сосна закрепляет их. Сосны выделяют особые летучие вещества, которые подавляют развитие многих вредных бактерий не только в лесу, но и в его окрестностях. Хвойные леса, как и лиственные, задерживают таяние снега, что обогащает почву влагой. В народном хозяйстве древесину сосны и ели используют как ценный строительный и поделочный материал. С помощью химической обработки из древесины сосны получают искусственные волокна, подобные шелковым нитям. Из древесины ели изготовляют бумагу. Древесина голосеменных растений — ценное сырье для многих отраслей промышленности. Сосну сибирскую называют в Сибири кедром. Но настоящие кедры растут в горах Северной Африки, на востоке Средиземноморской области и в Гималаях; у нас их культивируют на Черноморском побережье Кавказа и в Крыму. Из семян сосны сибирской получают хорошее пищевое «кедровое» масло.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1522; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.67.189 (0.013 с.) |