Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Блок 1 дополнительного иллюстративного и дидактического материала.
Характеристики рамет стабильны и мало отличаются от средних значений для соответствующего клона, что легко фиксируется визуально (рис. 4.14, 4.15).
Рис. 4.6. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 1 (архив клонов № 1) близкий к 600
Рис. 4.7. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 17 (архив клонов № 1) близкий к 700
Рис. 4.8. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 45 (архив клонов № 1) более 750
Рис. 4.9. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 22 (архив клонов № 1) более 700
Рис. 4.10. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 14 (архив клонов № 1) близкий к 900
Рис. 4.11. Угол крепления скелетных ветвей к стволу у клона плюсового дерева № 41 (архив клонов № 1) близкий к 900
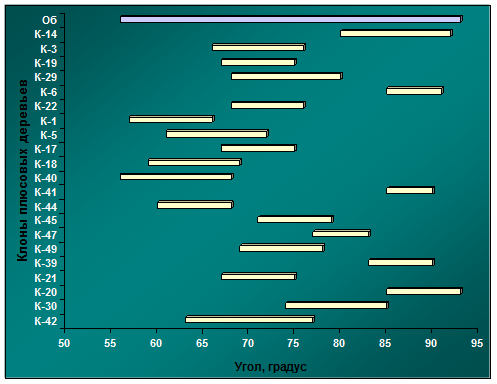
Рис. 4.12. Диапазоны изменчивости углов крепления скелетных ветвей к стволу в архиве клонов № 1
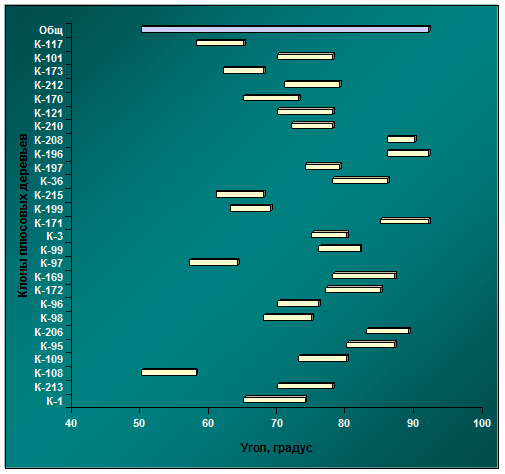
Рис. 4.13. Диапазоны изменчивости углов крепления скелетных ветвей к стволу в архиве клонов № 4
Рис. 4.14. Одинаковая величина угла крепления скелетных ветвей к стволу у рамет одного клона (плюсовое дерево № 1 в архиве клонов № 1) Рис. 4.15. Одинаковая величина угла крепления скелетных ветвей к стволу у рамет одного клона (плюсовое дерево № 29 в архиве клонов № 1) Оценить наличие существенных различий между изучаемыми клонами удалось с помощью дисперсионного анализа. Для этого исследованное вегетативное потомство плюсовых деревьев, достигших генеративного возраста, было объединено в неравномерный двухфакторный иерархический комплекс. Возможность построения обобщенного дисперсионного комплекса для всех изучаемых объектов (архивов клонов) обусловлена тем обстоятельством, что значения углов крепления скелетных ветвей к стволу являются хронологически стабильным признаком. Измерения на всех учетных деревьях осуществлялись в пределах одного возрастного диапазона заложения скелетных ветвей – до 10 лет (от нижних мутовок). Дисперсионный комплекс имел следующие характеристики: общее число наблюдений – 3711; число клоновых групп (градации по фактору высшей иерархии) – 48; число учетных деревьев (градации по фактору низшей иерархии) – 303. Числа степеней свободы соответственно составили: общее – 3710; по фактору высшей иерархии «а» – 47; по фактору низшей иерархии «b» – 255; по всем организованным факторам «x» – 302; по случайным факторам «z» – 3408. Кроме того, был сформирован однофакторный комплекс, градациями которого выступали группы рамет каждого плюсового дерева (ортета) в целом. Результаты по вегетативному потомству 48 плюсовых деревьев сосны обыкновенной, отобранных в лесхозах Нижегородской области, сосредоточенному в близлежащих архивах клонов, приведены ниже (табл. 4.9).
Таблица 4.9 Результаты дисперсионного анализа по величине угла крепления ветвей
Наличие существенных различий между сравниваемыми образцами выявил однофакторный дисперсионный анализ, в котором действующим фактором выступали различия в происхождении плюсовых деревьев (см. табл. 4.9). Для этого сравнивались между собой клоны в целом. Опытный критерий Фишера (Fоп = 2369,87) во много раз превышает табличное значение (F05 = 1,6). Вычисленные величины наименьшей существенной разности (НСР05 = 0,53) и D-критерия Тьюки (D = 0,96) показывают, что между большинством сравниваемых объектов различия достигают уровня, когда их можно отнести к существенным. При этом сила влияния фактора, вызывающего возникновение фиксируемых различий, оценивается как 0,9682±0,0004. Полученные результаты означают, что видимые различия в углах крепления ветвей между клонами плюсовых деревьев практически полностью определены их генетическими характеристиками. Факторы среды оказывают минимальное влияние на проявление данного признака, что вполне закономерно в условиях выровненного экофона и высоком уровне контроля со стороны генотипа сравниваемых объектов. Проведенный двухфакторный иерархический дисперсионный анализ, в котором фактором высшей иерархии выступали различия между клонами, а фактором низшей иерархии – различия между раметами, установил существенность различий между сравниваемыми объектами по комплексу всех организованных факторов (различия в происхождении плюсовых деревьев и различия между учетными деревьями). Расчетные критерии Фишера (Fx = 408,22) значительно превышают соответствующие табличные значения (F05 = 1,2 F01 = 1,3).
Предоставляемая иерархическим анализом возможность вскрыть структуру организованных факторов позволила оценить действенность каждого из них отдельно. Действие фактора высшей иерархии (различия между клонами) оказалось значимым. Оценка его дисперсионного отношения (Fa = 1063,48) заметно превышает табличный уровень (F05 = 1,4 F01 = 1,6) и позволяет признать наличие существенных различий между клонами. Это означает, что видимые различия между сравниваемыми объектами преимущественно обусловлены разницей в происхождении собственно плюсовых деревьев, разницей в их генотипах. Действие организованных факторов проявилось и в различиях между раметами, составляющими дисперсионный комплекс. Однако различия между ними не столь контрастны, хотя и являются существенными, а их оценки (Fb = 2,45) превышают критическую табличную величину (F05 = 1,2) на 5% уровне значимости и не превышают её на 1%.(F01 = 1,3). Оценивая величину влияния и степень существенности этих различий, мы исходили из того, что между раметами одного ортета не может быть генотипически обусловленных различий и, следовательно, природа зафиксированной неоднородности преимущественно определена разницей между представителями разных клонов. Расчет эффективности действия факторов, сделанный с учетом неравномерности анализируемого дисперсионного комплекса, позволил установить, что влияние различий в происхождении клонов (собственно различий между плюсовыми деревьями) доминирует (0,96). Поскольку в условиях опыта различия между клонами проявились на выровненном экофоне и обусловлены только разницей в происхождении самих плюсовых деревьев, то полученные данные свидетельствуют о высокой степени наследственной обусловленности различий по такому признаку как угол крепления ветвей к стволу. Сила влияния такого фактора как различия между отдельными учетными деревьями комплекса достаточно мала (0,01). Ограничено и влияние неорганизованных факторов (0,03). Это соответствует представлению о неорганизованных факторах, как о факторах, влияние которых рассматривается преимущественно как влияние различий в условиях среды, как экологическая составляющая общей фенотипической изменчивости. Кроме того, это соответствует представлениям о характере реакций представителей одного вида на экологический фон. У близких по происхождению объектов, имеющих сходные адаптационные характеристики, что имеет место в анализе, естественно ожидать и однотипные по характеру и масштабам реакции на природные условия территориально ограниченного участка, каковыми можно считать лесосеменные плантации, архивы клонов и другие объекты ЕГСК (ПЛСБ).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-06-14; просмотров: 67; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.69.45 (0.007 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||