Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Генетика и селекция растений. Генетика и селекция растенийСодержание книги
Поиск на нашем сайте
З.Г. Малышева ГЕНЕТИКА И СЕЛЕКЦИЯ РАСТЕНИЙ ПРАКТИКУМ для бакалавров, обучающихся по направлению «Лесное дело»
Новочеркасск 2018 Министерство сельского хозяйства Российской Федерации Новочеркасский инженерно-мелиоративный институт им. А.К. Кортунова ФГБОУ ВО «ДОНСКОЙ ГАУ»
З.Г. Малышева ГЕНЕТИКА И СЕЛЕКЦИЯ РАСТЕНИЙ ПРАКТИКУМ для бакалавров, обучающихся по направлению «Лесное дело»
Новочеркасск 2018
УДК 630*165.3 (075.8) М 207 Практикум составила д.-р с.-х. наук, профессор Малышева З.Г.
Рассмотрен на заседании кафедры лесоводства и лесных мелиораций (протокол № 6 от 16.01. 2018 г.) и рекомендован к изданию методическим советом лесохозяйственного факультета (протокол № 5 от 22.01.2018 г.)
Рецензент- Маркова И.С.-доц. каф.лесоводства и лесных мелиораций
Малышева З.Г. М 207 Генетика и селекция растений [Текст]: практикум для бакалавров, обучающихся по направл. «Лесное дело» / З.Г. Малышева; Новочерк. инж.- мелиор. ин-т ДГАУ.- Новочеркасск, 2018 - 136 с.
В практикуме изложены основные положения по наследственности и изменчивости древесных растений, хромосомная теория, и молекуляр-ные основы наследственности, закономерности наследования признаков при внутривидовой гибридизации, морфологические особенности строения пыльцы растений, техника скрещивания на срезанных ветвях, вегетативное размножение древесных и кустарниковых пород и расчет потребности привитого посадочного материала для создания клоновой лесосеменной плантации. Предназначено для помощи бакалаврам в освоении методики и техники селекционной работы с древесными и кустарниковыми растениями.
Ключевые слова: наследственность, изменчивость, ДНК, РНК, законы Менделя, гибридизация, отбор, семеноводство, плюсовые деревья, постоянная лесосеменная база. ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ Генетика и селекция растений является фундаментальной дисциплиной, формирующей мировоззрение в области биологии, почвоведения, ботаники, дендрологии и физиологии. Генетические закономерности лежат в основе всех биологических явлений, их познание и использование дают возможность управлять индивидуальным развитием и формообразованием, создавать высокопродуктивные породы растений. Целью настоящих методических указаний является формирование у студентов системы знаний по фундаментальным генетическим основам возникновения и функционирования живых организмов и биоценозов и приобретении навыков по вегетативному размножению древесных и кустарниковых растений. Для желающих более полно ознакомиться с современными достижениями в области генетики и селекции растений, а также с некоторыми разделами общей генетики в конце методических указаний дан список рекомендуемой литературы.
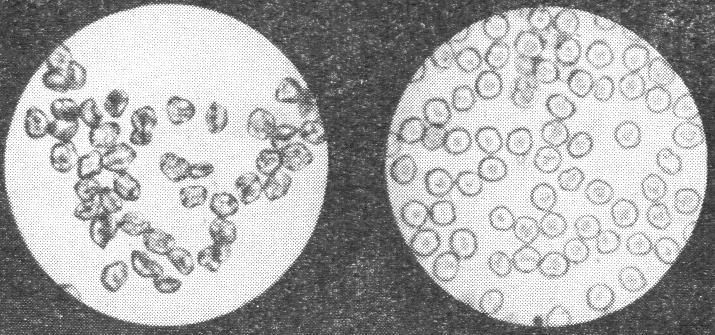
Форма и размеры клеток
б) Используя общие правила работы с биологическим микроскопом студенты рассматривают временный препарат с продольными срезами корешков под микроскопом при увеличении 15×, определяя форму и размеры клеток. Форма клеток определяется их функциями и местоположением в организме и как правило, имеют шаровидную или овальную форму. Есть клетки, не имеющие постоянной формы, она меняется в зависимости от выполняемых в разное время функций. Клетки, входящие в состав тканей и органов растений обычно значительно различаются по ширине и длине, часть вытянуты и имеют заостренные концы. Размеры клеток также разнообразны. Диаметр их колеблется от нескольких микрометров до нескольких сантиметров, длина – от 200 мкм до 1000 мм. Общая численность клеток в организме растения выражается огромными цифрами. Цвет клеток может быть от белого до черного, т.е содержит весь спектор окраса. На основании изучения препаратов выполняются их рисунки.
Законы Г. Менделя в) Студенты изучают законы Г.Менделя на основании приведенных данных, которые показывают, что гибриды F1 в соответствии с правилом единообразия все красноцветковые, но они образуют и яйцеклктки и спермии двух типов А и а. При оплодотворении на основе равновероятного сочетания двух типов гамет получается три типа зигот: АА, Аа и аа. Красная окраска цветков доминирует над белой, поэтому в F2 происходит расщепление в отношении 3 красноцветковых: 1 белоцветковое растение. Белоцветковые растения в F2 при дальнейшем размножении будут давать только белоцветковое потомство. Все они оказываются одинаковыми по фенотипу и по своей структуре (генотипу). Во время мейоза у гибридного растения F1 материнские хромосомы, несущие доминантный ген, и отцовские хромосомы, несущие рецессивный ген, расходятся в дочерние клетки независимо друг от друга, и поэтому при случайном соединении гамет во время оплодотворения образуется три типа зигот. Половина из них будет гибридной, ¼ воспроизведёт материнский тип и ¼ - отцовский. Красноцветковые растения F2 одинаковые по фенотипу, но различные по генотипу: 1/3 их имеет два одинаковых доминантных гена (АА), 2/3 – по одному доминантному и рецессивному гену (Аа). Гомозиготные особи (АА, аа) при размножении не дают расщепление в последующих поколениях, гетерозиготные формы (Аа) продолжают расщепляться. На основании вышеизложенного студенты зарисовывают порядок размножения гомо- и гетерозиготных форм в соответствии с рисунком 9.
Р Р ♀ F1 ♂
F2
Рисунок 9 – Размножение гомозиготных и гетерозиготных форм при моногибридном скрещивании
Из данной схемы видно, что при полном доминировании число классов гибридных организмов F2 по фенотипу и генотипу не совпадает т.к. при моногибридном скрещивании по фенотипу выделяется два класса: красноцветковые и белоцветковые особи, а по генотипу – три класса в отношении 1: 2: 1 (особи с генетической структурой АА, Аа, аа). Зная порядок размножения особей, студенты описывают поведение гомологичных хромосом при моногибридном скрещивании в соответствии с таблицей 5.
Таблица 5. Поведение пары гомологичных хромосом при моногибридномскрещивании
Из таблицы видно, что генотип в F2 имеет ¼ часть особей гомозиготных с доминантным признаком и ½ (2/4) гетерозиготных особей, имеющих в генах и доминантный и рецессивный признаки. Установленные Г. Менделем закономерности позволили сформировать два первых его закона: Первый закон – закон доминирования или единообразия гибридов первого поколения. Второй закон – закон определённого количественного расщепления гибридов второго поколения. Количественное расщепление по фенотипу при моногибридном скрещивании в F2 3: 1, а по генотипу 1: 2: 1.
При расщеплении Ход работы: а) Студенты дают определение полигибридному скрещиванию и анализируют поведение альтернативныхпризнаеков при расщеплении в F2 по генотипу и фенотипу. Скрещивание особей, различающихся по трём и более парам аллельных признаков, называются полигибридными. В общем виде полигибридное скрещивание можно представить следующим образом: Р ААВВССDD × ааввссdd F1 АаВвСсDd F2 (А + а)2 × (В + в)2 × (С + с)2 × (D + d)2 Эти признаки дают более сложную картину расщепления по сравнению с дигибридными скрещиваниями, подчиняются тем же закономерностям наследования. Анализ наследования одной пары признаков в моногибридном скрещивании позволяет понять независимое наследование двух и более признаков при дигибридном и полигибридном скрещивании (три, четыре и более генов). Расщепление в F2 по фенотипу для каждой пары альтернативных признаков равно 3: 1. Это исходное отношение обеспечивается точным цитологическим механизмом расхождения хромосом в мейозе. Принцип независимого поведения разных пар альтернативных признаков в расщеплении по фенотипу в F2 выражается формулой (3: 1)n, где n – число пар альтернативных признаков. Задачи 1. Какие типы гамет образуют растения, имеющие генотипы: а) АаВВСс; б) ааВвСс; в) АаВвСс. У гороха гладкая форма семян А доминирует над морщинистой а, жёлтая окраска семян В над зелёной в и красная окраска цветков С над белой с. 2. Скрестили гомозиготные растения, отличающиеся по четырем парам признаков. Определить: а) число и соотношение классов гибридных особей в F2 по фенотипу; б) число классов гибридных особей в F2 по генотипу. 3. ГетерозиготаАаВвСсDd скрещена с гомозиготным рецессивом. Определить: а) число классов в полученном потомстве по генотипу; б) какая часть потомства имеет все четыре доминантных гена; в) какая часть потомства имеет все четыре рецессивных гена. 4. Произведено гексагибридное скрещивание. Определить число классов по генотипу и фенотипу в F2.
Популяция и ее виды Ход работы:
а) Студенты дают определение популяции и ее видам. Популяция – совокупность особей одного вида, заселяющих определённую территорию, свободно скрещивающихся друг с другом и в той или иной степени изолированных от других совокупностей. В практической селекции под популяцией понимают группу особей, имеющих наследственные различия. Популяции могут быть следующих видов: Популяция гибридная – совокупность наследственно различающихся растений, полученная в результате гибридизации. Популяция естественная – сформировавшаяся под действием естественных факторов. Популяция идеальная – не существующая в природе популяция, параметры которой используются только в математических расчётах популяционной генетики. Такая популяция характеризуется бесконечно большим постоянно сохраняющимся объёмом, панмиксией, свободной от мутаций, действия отбора и внешней среды. Реальность идеальной популяции исключается, поскольку в противном случае исключалась бы эволюция. Популяция изогенная – все особи которой генетически идентичны в отношении определённых генов, т.е. все особи или гомозиготны по этим генам, или все гетерозиготны по ним. Популяция изогенная является следствием инцухта (инбридинга) – это развитие в себе, скрещивание особей, родство между которыми более тесное, чем родство между особями, случайно взятыми из той же популяции. У перекрёстноопыляемых растений – это принудительное самоопыление, повторяющиеся в большом или меньшем числе последовательных поколений, что ведёт к возрастанию гомозиготности и в следствии гомозиготации получают инцухт – линии, называемых также инбредными или самоопылёнными. Популяция менделевская – совокупность в определённой степени сходных по фенотипу и генотипу особей, заселяющих определённый географический ареал, характеризующихся одинаковым способом размножения, одинаковой наследственной изменчивостью и подверженностью действию отбора и внешней среды. Крайним пределом объёма такой популяции является биологический вид. Все виды организмов состоят из популяций. ВИД – это основная систематическая единица, реально существующая в природе, занимающая определённый ареал и представляющая совокупность родственных по происхождению особей, качественно отличных от других видов и генов скрещивающихся с ними. Популяция – это главный структурный элемент вида, форма его существования в данных условиях. На основе генетических преобразований в популяции идут микроэволюционные процессы, завершающиеся видообразованием. Вид постепенно складывается и приспосабливается отбором к определённым экологическим условиям на основе генетических преобразований, происходящих в популяциях часто с такой быстротой, что их нельзя объяснить действием индивидуального отбора. Приспособление вида к меняющимся условиям среды не сводится к морфологическим и физиологическим изменениям отдельных особей или их признаков. В эволюции и селекции происходит не просто изменение признаков и свойств организмов, а идёт замена одной нормы реакции генотипа популяции другой. Вид представляет собой генетически закрытую систему, а популяция – система генетического открытия. Поэтому, процесс видообразования в общей форме сводится к преобразованию генетически открытых систем в генетически закрытые. Несмотря на дискретное строение наследственных единиц, изменчивость организмов имеет непрерывный характер, а эволюционный процесс принципиально безграничен. Популяция является его элементарной единицей. Эволюция организмов совершается путём непрерывной замены в популяции генотипов другими. Генетическая изменчивость складывается из мутационной и комбинационной изменчивости. Каждая популяция имеет определённый генофонд, генетическую структуру, связанную с составом хромосомного набора и относительным качеством разных генов. Генетическая структура популяции определяет её свойства. На формирование и обособление популяций, их структуру оказывают влияние многие факторы: интенсивность и направление отбора, способ размножения, миграции, характер и темп мутационной изменчивости, численность особей, различные виды изоляции и др., главным из них является отбор, а также способ размножения. Популяция – это единая система обмена генетической информацией между составляющими её особями. С. С. Четверяков отмечал, что вид, как губка, впитывает гетерозиготныегеновариации, оставаясь при этом внешне (фенотипически) однородным. За внешним однообразием популяции скрыто огромное разнообразие её генетического материала. Благодаря этому популяция приобретает высокую пластичность для приспособления к постоянно меняющимся условиям существования. Закон Харди – Вайнберга
б) Студенты изучают генетические процессы в популяциях на основании Закона Харди-Вайнберга, который гласит, что если популяция находится в равновесии, то частота генотипов и генов у родителей и потомства не меняется. Под "равновесием" понимается отсутствие изменений частот генотипов в последовательных поколениях одной популяции. Особое состояние равновесия описывается с помощью закона или правила Г. Харди-Вайнберга (математик Г. Харди и немецкий врач Н. Вайнберг) которому подчиняется частота распределения гетерозигот в свободно скрещивающейся популяции, и выразили его в виде алгебраической формулы. Оказалось, что частота членов пары аллельных генов в популяции распределяется в соответствии с коэффициентом разложения бинома Ньютона (P+g)2. Закон Харди – Вайнберга выражает вероятностные распределения генотипов в любой свободно скрещивающейся популяции. Но действие этого закона предполагает выполнение ряда обязательных условий: 1) популяция имеет неограниченно большую численность; 2) все особи в популяции могут совершенно свободно скрещиваться; 3) гомозиготные и гетерозиготные по данной паре аллелей особи одинаково плодовиты, жизнеспособны и не подвергаются отбору; 4) прямые и обратные мутации происходят с одинаковой частотой или они так редки, что ими можно пренебречь. Совершенно очевидно, что все эти условия в реально существующих популяциях не выполнимы, и закономерности, установленные Харди и Вайнбергом, правдивы только для идеальной популяции. Но этот закон является основой для анализа динамики генетических преобразований, совершающихся в реальных естественных популяциях при нарушениях, вызванных действием эволюционных факторов: отбора при возникновении мутаций; ограничений численности особей и т.д.
Адаптация и ее виды
Ход работы:
а) Студенты дают определение понятию адаптация, под которой понимается процесс любых изменений в структуре или функциях организма, обеспечивающих способность к существованию. При этом устойчивость организма зависит от их адаптации к окружающей среде. Различают понятия адаптация и адаптивность. Адаптация – это способность организмов жить и размножаться в данных условиях среды. Адаптивность – это способность к адаптации (приспособлению) в определённых границах условий внешней среды. Те структурные изменения организма, которые увеличивают его жизнеспособность, выживаемость, темп размножения, называются адаптивными признаками. Организмы на протяжении всей своей жизни приобретают определённую норму реакции на изменение условий окружающей среды. В связи с этим различают адаптацию генотипическую и модификационную. Генотипическая адаптация – это адаптация, при которой изменение генотипа обеспечивает образование новой нормы реакции и гармоничное приспособление индивида к новым экологическим условиям. Модификационная адаптация – это адаптация, позволяющая организму в пределах сложившейся нормы реакции оставаться жизнеспособным и продуцировать потомство в новых условиях среды. Кроме этого различают онтогенетическую адаптацию, которая характеризует приспособительные изменения в период индивидуального развития организма. Филогенетическая адаптация – это результат естественного отбора в ряде поколений.
Основные положения:
Заготовка пыльцы. Зрелые сережки (единичные пыльники начинают лопаться и из них высыпается пыльца) срезают ножницами с ветвей и собирают в бумажные (целлофановые) пакеты. Затем переносят в лабораторию. Здесь сережки раскладывают тонким слоем (в одну сережку) на листе чисто глянцевой бумаги, при этом все примеси (частички листьев, почечных чешуй и т.п.) удаляют. Затем листы с разложенными сережками помещают в затемненном месте. Если сережки собраны вполне зрелыми, то через 1-2 часа пыльники, подсохнув, начинают лопаться и пыльца в массе высыпается на бумагу. После этого пинцетом удаляют сережки, но на поверхности остается еще много примесей, которые путем поднятия листа и легкого постукивания пальцами по нему стараются стряхнуть все примеси с поверхности пыльцы. После этого чистую пыльцу собирают в центре листаиосторожно ссыпают в чистую тару, на которую наклеивается этикетка с полным указанием породы, место сбора, дата сбора, номер партии, если собрано несколько партий, все эти сведения заносятся в «Журнал пыльцы» (таблица 10). Приготовленную этикетку вкладывают в сосуд с пыльцой, закрывают и помещают в эксикатор и в холодильник. Для сохраненияжизнеспособности пыльцы необходимо чтобы температура была близкой к 00С. Таблица 10 – Журнал пыльцы
Общая морфология пыльцы древесных пород
Чтобы правильно описать морфологию пыльцы, необходимо предварительно ознакомиться с общей морфологией пыльцы древесных пород. Пыльцевые зерна имеют 2 оболочки: наружную, кутинизированную – экзину и внутреннюю, нектиновую – энтину. Толщина этих оболочек различна, причем имеется некоторая зависимость между их размерами: если одна из них имеет большую толщину, то другая, наоборот, бывает более тонкой. Интина состоит из светлого стекловидного вещества, способного сильно набухать в воде. Экзина состоит из нескольких слоев (чаще из 3). Она может быть как совершенно гладкой, так и покрытой выростами и углублениями различной величины и формы, т.е. имеет скульптуру. Чаще всего пыльца ветроопыляемых пород бывает гладкой или со слабовыраженной скульптурой. Пыльца насекомоопыляемых видов имеет рельефные скульптурные образования: шипы, бугорки, ямки и т.п. На поверхности пыльцевого зерна следует еще различать борозды и поры. Борозды – это продольно-вытянутое углубление в экзине, покрытое более тонким ее слоем, так называемой мембраной. Борозда является местом прорастания пыльцевой трубки и регулятором объема зерна в зависимости от степени его набухания, поэтому при набухании зерна края борозды раздвигаются и мембрана оказывается натянутой. По форме борозды могут быть различны от длинных до округлых. Количество борозд, в зависимости от вида, колеблется от 1 до 30 и более. У покрытосеменных, двудомных чаще из 3, расположены они как бы меридиально на равных расстояниях одна от другой. Оси борозд сходятся у полюсов под углом 1200. У голосеменных пыльцевых зерен имеется всего одна борозда, желобчатой формы, помещающаяся на внешней, углубленной стороне зерна. Поры – это отверстия в экзине, из которых выходит пыльцевая трубка. Количество пор и их форма разнообразны и обычно являются постоянными для пыльцевых зерен определенных систематических групп. Число их колеблется от 1 до 30 и более. Поры часто располагаются в бороздах и в определенном порядке, характерным для каждого вида. У голосеменных пород поры встречаются редко, однако они имеют воздушные мешки – выросты экзины, чаще их бывает 2. Располагаются воздушные мешки по бокам пыльцевого зерна или часто сдвинуты с наружной его части и размещены по краям борозды. По своим очертаниям воздушные мешки бывают полушаровидные или более чем полушаровидные. Ниже приводится описание пыльцы древесных пород. Голосеменные: Род Pinus – сосна. Пыльцевые зерна большие (длина 60-105 микрон, высота тела 30-60 мк, высота воздушных мешков 24-50 мк). Зерна состоят из тела и двух воздушных мешков, расположенных по краям слабоочерченной борозды. Пыльца обладает исключительной летучестью и производится в огромном количестве. Экзина зернистая. У сосны обыкновенной мешки правильной полушаровидной формы, резко отделяются от тела зерна, линия прикрепления мешков к телу менее их диаметра. Воздушные мешки смещены к внутренней стороне пыльцевого зерна. Покрытосеменные: Род Salix – ива. Пыльцевые зерна в сухом состоянии эллипсоидальные, сильно вытянутые, в набухшем почти округлые. Пыльца 3-х борозчатая, без пор. Борозды меридиональные, доходят почти до полюсов, делят пыльцевое зерно как бы на 3 лопасти, что хорошо видно при наблюдении «с полюса». Экзина сетчато-ячеистая. Часто ячеи-сетки имеют перегородки, образующие шишкообразные выступы. Диаметр пыльцевых зерен 15-28 микрон. Род Populus – тополь. Очертания зерен округлые, экзинатонкая, гладкая или слабосетчатая. Борозд и пор нет. Диаметр 24-38 мк. Род Juglans – орех. Сухая пыльца сплющенно-сфероидальной формы вследствие того, что на одной из сторон зерна имеется участок утонченной экзины, вдавленной внутрь зерна. При набухании этот участок расправляется и зерно принимает вид почти правильного шара или многогранника с большим количеством углов, каждому углу соответствует пора, их от 8 до 16 штук. Поры расположены по экватору. Экзинатолстая, лишенная скульптуры или грубозернистая. Диаметр зерен 24-50 микрон. Род betulaceae – береза. Зерна эллиптические при рассмотрении сбокуи треугольно-округлые сверху («с полюса»). Пор – 3, редко 4 или 6. экзинагладкая, вокруг пор сильноутолщенная. Диаметр зерен 15-25 микрон. Род Corylus – орешник. Пыльцевые зерна при рассмотрении сбоку овальные, сверху округло-треугольные. Экзина гладкая. Зерна имеют 3 поры, которые расположены по углам треугольника. Экзина вокруг пор незначительно утолщена. Диаметр зерен 22-28 микрон. Род Quercus – дуб. Сухая пыльца - эллиптическая, при рассмотрении сверху – трехлопастная. Три борозды расположены меридиально, но не вполне достигают полюсов, с суженными концами. Поры отсутствуют. Экзина толстая, грубая, не утончающаяся к бороздам, скульптура зернистая. У различных видов дуба пыльца отличается только размерами и крупностью скульптуры. Диаметр зерен 16-36 микрон. Род Ulmus – ильм, вяз, берест, карагач. Пыльцевые зерна сплющенно-сфероидальной формы, без борозд, пор от 3 до 12, расположены по экватору, что обуславливает угловатость пыльцевых зерен при рассматривании их с «полюса» (рисунок 13). Чаще всего пор 5, вследствие чего пыльца имеетокругло-пятиугольное очертание. Скульптура в виде рельефных выпуклостей, напоминающих по рисунку извилины головного мозга. Размеры зерен: у береста 27-39 микрон, у вяза 24-36 мк, у карагача 30-50 мк.
М 1: 0,01 мм Рисунок 13 – Сухая (слева) и набухшая (справа) пыльца Ulmus (ориг). Род Acer – клен. Зерна в набухшем состоянии округлые, борозд 3 или 4, иногда 2 у клена ясенелистного. Некоторые виды кленов имеют поры. Борозды глубокие и широкие, с толстой мембраной. Если имеются поры, то они располагаются по одной в каждой борозде. Поверхность экзины покрыта тонкими то более, то менее ясно выраженными линиями, расположенными рядами различной длины. Размеры зерен у клена остролистного 30-40 микрон, у клена ясенелистного 27-37 микрон. Род Tilia – липа. Зерна чечевицеобразной формы с 3 или 4 порами, расположенными по экватору на равных расстояниях одна от другой в небольших углублениях. Поверхность экзины испещрена мелкими ямками, которые придают контурам зерна извилистый характер. Размер зерна 27-35 микрон. Род Fraxinus – ясень. Пыльцевые зерна имеют 3-4 иногда 5 борозд. Сверху пыльцевые зерна треугольные, четырехугольные или пятиугольные в зависимости от числа борозд, сбоку – эллиптические. Экзина ячеистая или бугорчатая. Размеры: длина 16-27 мк, ширина 19-22 мк. Ход работы:
Порядок работы при просмотре пыльцы:
Общие очертания пыльцевых зерен зависят от степени их набухания: сухая пыльца обычно имеет продолговато-эллиптическую форму, а набух-шая – округлую или шаровидную. 1) Просмотр пыльцы в сухом состоянии производится так: на чистое предметное стекло при помощи препаровальной иглы наносят небольшое количество пыльцы и рассматривают ее под микроскопом вначале под небольшим увеличением, а затем при увеличении 280х (объектив 40, окуляр 7х). Измеряют длину и ширину пыльцевых зерен при помощи окуляр-микрометра, результаты измерений записывают в тетрадь. Затем делают зарисовки формы пыльцевого зерна и описывают морфологические особенности пыльцевых зерен рассматриваемой породы. 2) Просмотр пыльцы в набухшем состоянии. Измерение набухшей пыльцы, не имеющей воздушных мешков, делают по двум взаимно перпендикулярным направлениям: наибольшей и наименьшей. Пыльцу с воздушными мешками измеряют в 3-х направлениях: 1) общая длина зерна, высота тела зерна, высота воздушных мешков. Измерение делают при увеличении 230х. В этом случае на предметное стекло наносится капля воды с небольшим количеством пыльцевых зерен, препарат накрывают покровным стеклом и под микроскопом изучают форму пыльцевых зерен в набухшем состоянии. Опять же выполняют замеры, зарисовывают и описывают морфологию набухших зерен рядом с рисунками. Морфологическое описание пыльцы ведется по следующей программе: общие очертания сухой и набухшей пыльцы, наличие пыльцевых мешков, борозд и пор, их количество и расположение, если есть воздушные мешки, то описывается, как они прикрепляются к телу пыльцевого зерна и какую форму (очертания) имеют; скульптуру экзины; толщина экзины и интины; средние размеры пыльцевых зерен (в сухом и набухшем состоянии) по длине и ширине, колебания этих размеров от максимума до минимума. Размеры пыльцевых зерен указываются в микронах, для чего полученные результаты измерения в делениях окуляр-микрометра умножают на цену деления при различном увеличении микроскопа. После изучения пыльцевых зерен одной породы, приступают к изучению следующего вида и в той же последовательности.
ПОДБОР ПОРОД, СОСТАВЛЕНИЕ СХЕМ СМЕШЕНИЯ, РАЗМЕЩЕНИЯ РАСТЕНИЙ И РАСЧЕТ ПОТРЕБНОСТИ ПОСАДОЧНОГО МАТЕРИАЛА ДЛЯ СОЗДАНИЯ ФИЛЬТРУЮЩЕЙ ЛЕСНОЙ ПОЛОСЫ ВОКРУГ МАТОЧНОЙ ПЛАНТАЦИИ
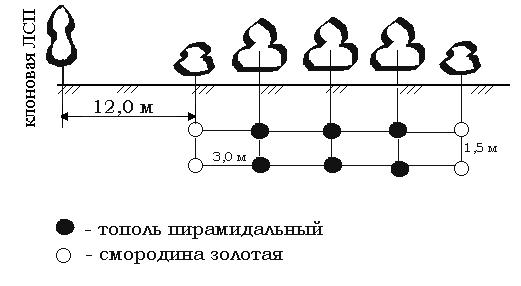
Цель работы: создание защитной лесной полосы и расчет потребности и стоимости посадочного материала для создания фильтрующей лесной полосы. Задачи: уметь решать задачи по определению потребного количества посадочного материала для создания лесных полос. Общие положения: для снижения заноса фоновой пыльцы по периметру ЛСП рекомендуется создание фильтрующих защитных полос из 5-10 рядов быстрорастущих, густокронных деревьев других видов, не являющихся промежуточными хозяевами опасных вредителей и грибных болезней. Для расчета потребности количества посадочного материала для создания лесной полосы следует составить схему лесной полосы, например 5-ти рядной, рисунок 17. Выбор схемы размещения растений в лесной полосе обуславливается биологическими особенностями видов (быстрота роста надземной части, необходимая площадь ризосферы и т.п.).
Рисунок 17 – Поперечный профиль лесной полосы
В лесную полосу рекомендуется вводить в крайние ряды кустарники для создания плотной конструкции. В нашем примере расстояние между деревьями в ряду 1,5 м и в междурядьях 3,0 м.
Ход работы:
Расчет потребности посадочного материала ведем по формуле:
где N – общее количество сеянцев одной породы в лесной полосе, шт.; L – протяженность (периметр) лесной полосы, м n – количество рядов в лесной полосе, шт.; l – расстояние между сеянцами в ряду.
Количество потребного количества посадочного материала рассчитывается по каждой породе отдельно с учетом отпада, данные заносим в таблица 16.
Таблица 16 – Количество посадочного материала, необходимого для создания защитной лесной полосы.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-06-14; просмотров: 112; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.119.255.170 (0.02 с.) |