Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиология растений как наука. Цели и задачи. Методы исследования. Значение для учителя.Стр 1 из 31Следующая ⇒
Физические и коллоидные свойства цитоплазмы,ее строение. По хим. Составу состоит из80% воды, остальное сух. -во ¾ белков..Цитопл-ма- коллоидный раствор белков,кот. способен к коагуляции и коацервации. Все белки имеют заряд и окружены гидратной оболочкой. Легко скользят образуя движение-такое состояние-ЗОЛЬ. Если уменьшается содержание воды в гидратных оболочках, белки слипаются образуя коацерваты. Свойства: способность вытягиваться в тонкие длинные нити, раздражимость, движение(колебательные, струйчатое). Белки цитоплазмы благодаря разнообразию их строения, химической природы, гетерополярности могут вступать в безграничное количество реакций с различными веществами, которые содержатся в цитоплазме или поступают извне. В результате этих реакций может измениться форма макромолекулы, что приведет к изменению ее химической активности. Таким образом, изменчивость свойств белков — важная особенность живого вещества. Цитоплазма построена по коацерватному типу и представляет сложную коллоидную систему из белковых, углеводных и липидных соединений. Белки относятся к гидрофильным коллоидам. белки обладают способностью денатурироваться. При денатурации гидрофильные коллоиды — белки — становятся гидрофобными, теряют стойкость и вследствие этого легко коагулируют. Важную роль во всех этих процессах играет поверхность цитоплазмы: она является средой для осуществления процессов адсорбции и десорбции, что влияет на движение частиц, которое может иметь большую скорость, проходить одновременно в противоположных направлениях и влиять также на свойства самой цитоплазмы — вязкость, эластичность, проницаемость и др. Особенности цитоплазмы не позволяют рассматривать ее как истинно золеобразную жидкость, поскольку она по упругости приближается к гелю. Явление взаимного превращения золя в гель наблюдается на протяжении всей жизнедеятельности клетки. На состояние цитоплазмы влияют концентрация водородных ионов, а также соотношение между содержанием одно-и двухвалентных катионов. В присутствии кальция коагуляция белков в цитоплазме происходит при более низкой температуре. Свойства цитоплазмы обусловливаются сложностью многофазной, полидисперсной, коллоидной системы. Цитоплазма имеет три слоя: внешний — плазмалемма, внутренний — тоно-пласт и лежащий между ними — мезоплазма. Пограничные слои плазмалеммы и тонопласт вязкие и эластичные, а мезо-плазма более текучая и менее эластичная.
Таким образом, цитоплазма — это сложная гетерогенная коллоидная структура, которая включает большое количество различных компонентов. Дисперсной средой является комплексный гидрозоль с высоким содержанием белковых и других макромолекул, сахаров, неорганических солей, например фосфатов. Важную роль играет вода, которая насыщает всю систему коллоидов цитоплазмы, образуя непрерывную фазу. В живой цитоплазме постоянно происходят процессы новообразования и распада различных веществ, коагуляция коллоидов и их обратное превращение в золи, образование коацерватов, гелей и т.д. Эти процессы непосредственно зависят от состояния и свойств структур, из которых она состоит. Изменения протоплазменных структур под воздействием внешних условий имеют приспособительный характер. Следовательно, цитоплазма — система многофазная, подвижная, динамичная; она закономерно изменяется под влиянием внутренних и внешних факторов. В цитоплазме осуществляются метаболические циклы веществ и энергии, составляющие основу жизни растительного организма. Строение и физиологическая роль фикобиллинов. Содержание отдельных групп пигментов у растений разных мест обитания. Теория хроматической адаптации. Синезеленые водоросли (цианобактерии), красные морские водоросли и некоторые морские криптомонады помимо хлорофилла а и каротиноидов содержат пигменты фикобилины. Наиболее известные представители фикобилинов — фикоэритробилины и фикоцианобилины. Первые преобладают у красных водорослей и определяют их цвет, вторые — у синезеленых. Структура и свойства фикобилинов. По структуре фикобилины. (от греч. «phycos» — водоросль и лат. bilis — желчь) относятся к группе желчных пигментов — билинов (у животных представитель этой группы — билирубин). Это тетрапирролы с открытой цепью, имеющие систему конъюгированных двойных и одинарных связей. В своем составе они не содержат атомов магния или других металлов, а также фитола.
Значение фикобилинов. Максимумы поглощения света у фикобилинов находятся между двумя максимумами поглощения у хлорофилла: в оранжевой, желтой и зеленой частях спектра (см. рис. 4.3). Значение такого распределения максимумов поглощения становится понятным, если вспомнить оптические свойства воды, которая поглощает прежде всего длинноволновые лучи. На глубине 34 м в морях и океанах полностью исчезают красные лучи, на глубине 177 м — желтые, на глубине 322 м — зеленые и, наконец, на глубину свыше 500 м не проникают даже синие и фиолетовые лучи. В связи с таким изменением качественного состава света в верхних слоях морей и океанов обитают преимущественно зеленые водоросли, глубже — сине-зеленые и еще глубже — водоросли с красной окраской. В. Т. Энгельман назвал это явление хроматической комплементарной адаптацией водорослей. По его наблюдениям (1881 — 1884), наиболее интенсивная ассимиляция С02 у водорослей с различной окраской соответствует максимумам поглощения света пигментными системами этих водорослей. Русский исследователь Н. М. Гайдуков (1903) экспериментально показал, что если культуру синезеленой водоросли Oscillaria sancta выращивать на свету разного спектрального состава, то у нее развивается дополнительная (комплементарная) окраска. При освещении зеленым светом водоросли становятся оранжево-красными, а при дейтвии красных лучей — зелеными. В настоящее время известно, что эти изменения цвета клеток связаны с изменениями в синтезе фикобилинов, принимающих участие в процессе фотосинтеза. Таким образом, у водорослей фикобилины — дополнительные пигменты, выполняющие вместо хлорофилла b функции светособирающего комплекса. Около 90% энергии света, поглощенного фикобилинами, передается на хлорофиллы а. Явление хроматической комплементарной адаптации обнаружено далеко не у всех видов синезеленых и красных водорослей. У многих из них адаптация к изменяющемуся спектральному составу света обеспечивается изменением количества и состава хлорофиллов Пигменты — это окрашенные вещества. Пигменты поглощают свет определенной длины волны. Непоглощенные участки солнечного спектра отражаются, что и обусловливает окраску пигментов. Так, зеленый пигмент хлорофилл поглощает красные и синие лучи, тогда как зеленые лучи в основном отражаются. Видимая часть солнечного спектра включает длины волн от 400 до 700 нм. Вещества, поглощающие весь видимый участок спектра, кажутся черными. Состав пигментов зависит от систематического положения группы организмов. У фотосинтезирующих бактерий и водорослей пигментный состав очень разнообразен (хлорофиллы, бактериохлорофиллы, бактериородопсин, каротиноиды, фикобилины). Их набор и соотношение специфичны для различных групп и во многом зависят от среды обитания организмов. Пигменты фотосинтеза у высших растений значительно менее разнообразны. Пигменты, сконцентрированные в пластидах, можно разделить на три группы: хлорофиллы, каротиноиды, фикобилины. 11. Локализация пигментов в листе. Их связь с мембранами. Понятие о фотосинтетических единицах и фотосистемах.ФС1 и ФС2.. Пигменты растений, участвующие в фотосинтезе, "упакованы" в тилакоиды хлоропластов в виде функциональных фотосинтетических единиц, называемых фотосистемами. Имеется два типа фотосистем – I и II. Каждая фотосистема содержит 250-400 молекул пигментов. Все пигменты фотосистемы могут поглощать частицы световой энергии, называемые фотонами или квантами света, но только одна молекула хлорофилла данной фотосистемы может использовать поглощенную энергию в фотохимических реакциях. Эта молекула называется реакционным центром фотосистемы, а другие молекулы пигментов называются антенными, поскольку они улавливают энергию света, подобно антеннам, для последующей передачи реакционному центру.
Реакционные центры. Белки, содержащие длинноволновые формы хлорофиллов (П680 в ФС II и П700 в ФС I) в количестве одна молекула на 200 — 400 молекул других хлорофиллов и способные к первичному фотохимическому разделению зарядов, называют реакционными центрами В фотосистеме I реакционный центр образован особой молекулой хлорофилла a, обозначаемой как P700, где 700 – оптимум поглощения в нм. Реакционный центр фотосистемы II также образован молекулой хлорофилла a и обозначается индексом P680, поскольку оптимум поглощения лежит в районе 680 нм. Фотосистемы I и II работают обычно синхронно и непрерывно, но фотосистема I может функционировать отдельно. В эволюционном отношении ФС-1 более древняя фотосистема. У фотосинтезирующих бактерий эта система не способна использовать воду в качестве донора электронов и является единственной.ФС-1содержит примерно 200 молекул хлорофилла.В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). С-2 содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр ФС-2 представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофиллаaс максимумом поглощения при 680 нм (П680) феофитин. На него,в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680становится сильным восстановителем (E0=-0,7 В). Синтез АТФ Синтез АТФ осуществляется тремя способами: фотосинтетическое фосфорилирование, окислительное фосфорилирование (сопряжённое с транспортом электронов по дыхательной цепи) и субстратное фосфорилирование. В первых двух процессах преобразование поступившей с потоком электронов энергии в энергию фосфоэфирных связей АТФ осуществляет особый фермент — АТФ-синтетаза. Этот фермент присутствует во всех мембранах, участвующих в преобразовании энергии (мембраны бактерий, митохондрий и хлоропластов). АТФ-синтетаза катализирует присоединение неорганического фосфата (Фн) к АДФ, образование которого осуществляет аде-нилаткиназа
(АМФ + АТФ = 2 АДФ). Активность АТФ-синтетазы можно обнаружить по обратной реакции гидролиза АТФ: АТФ + Н20 = АДФ + Фн + Н+. Благодаря обратимости реакции фосфорилирования, накопившийся АТФ может быть использован для создания протонного градиента, обеспечивающего энергией движение жгутиков и осмотическую работу. Энергия также направляется для обратного переноса электронов, необходимого для восстановления никотинамидадениндинуклеотида (НАД) при использовании бактериями неорганических доноров электронов (S03, N03, Fe2+ и др.). Дыхательная электронтранспортная цепь (ЭТЦ) — система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану. Дыхательная электронтранспортная цепь состоит из переносчиков электронов, которые передают электроны от субстратов на кислород. Расположение переносчиков определяется величиной их окислительно-восстановительного потенциала. Таким образом, транспорт электронов в дыхательной электронтранспортной цепи сопровождается трансмембранным переносом протонов. Возникающая разность потенциалов по обеим сторонам внутренней мембране митохондрий используется для синтеза АТФ (окислительное фосфорилирование). В результате прохождения двух электронов по цепи образуется 3 молекулы АТФ. Компоненты ЭТЦ: НАД(Ф) – зависимые дегидрогеназы, растворимые ферменты; Белки – содержащие железоероцентры, изменение валентности; Хиноны – жирорастворимые соединения, имеющие длинный терпеноидный хвост, связанный с хиноидным ядром, наиболее распространен убихинон; цитохромы – переносят только электроны за счет изменения валентности железа из +2 в +3; колплекс цитохромов – цитохромоксидаза, присутствие Си вместо железа. Дыхательная цепь митохондрий содержит 4 основных комплекса и два небольших по молекулярной массе компонента – убихинон и цитохром. Комплекс 1 осуществляет перенос электронов от НАДН к убихинону. Комплекс 2 катализирует окисление сукцината убихиноном. Комплекс 3 переносит электроны от восстановленного убихинона к цитохрому. В комплексе 4 электроны переносятся от цитохрома к кислороды. Значение для фотосинтеза различных участков солнечного спектра. Образование углеводов и аминокислот при фотосинтезе.
Согласно первому закону фотохимии, только поглощённые лучи могут быть использованы в химических реакциях. В том случае, если реагирующие молекулы бесцветны и не поглощают свет, фотохимические реакции могут идти только в присутствии специальных веществ – фотосенсибилизаторов. Фотосенсибилизаторы – вещества, поглощающие энергию света и передающие её той или иной бесцветной молекуле. Положение о том, что в процессе фотосинтеза могут быть использованы только поглощённые лучи, впервые получило экспериментальное подтверждение в опытах К.А.Тимирязева. До этого господствовало ошибочное представление, что наибольшее значение в процессе фотосинтеза имеют жёлтые лучи солнечного спектра, которые хлорофиллом не поглощаются. К.А.Тимирязев показал, что процесс усвоения CO2 на свету представляет собой фотохимический процесс и подчиняется законам фотохимии, что процесс фотосинтеза проходит именно в тех лучах, которые поглощаются хлорофиллом. Хлорофилл является оптическим сенсибилизатором, поглощающим энергию света. Наиболее интенсивное усвоение углекислого газа наблюдается в красных лучах. Затем, в направлении зелёной части спектра процесс фотосинтеза постепенно ослабевает, зелёные лучи хлорофиллом почти не поглощаются. В сине-фиолетовой части спектра наблюдается второй подъём интенсивности фотосинтеза. Таким образом, кривая интенсивности фотосинтеза имеет два максимума соответственно двум максимумам поглощения хлорофилла. Интенсивность процесса фотосинтеза в различных участках спектра получила название спектра действия. Фотосинтез — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов. Световая фаза — это этап, на котором энергия света, поглощенная хлорофиллом, преобразуется в химическую энергию АТФ и НАДФН2. Осуществляется на свету в мембранах гран при участии белков-переносчиков и АТФ-синтетазы. Реакции, вызываемые светом, происходят на фотосинтетических мембранах гран хлоропластов: возбуждение электронов хлорофилла квантами света и их переход на более высокий энергетический уровень; Результатами световых реакций являются: образование кислорода, синтез АТФ, восстановление НАДФН2. Темновая фаза — процесс преобразования СO2 в глюкозу в строме хлоропластов с использованием энергии АТФ и НАДФН2. Реакции фиксации углерода — это последовательные преобразования СO2 в глюкозу: помимо молекул глюкозы в строме образуются аминокислоты, нуклеотиды, спирты. Углеводы являются важнейшей составной частью всех растений. Они образуются в зеленых растений в результате фотосинтеза из углекислого газа и воды под действием световой энергии. При этом углерод из неорганической формы переходит в органическую. По современным представлениям фотосинтез представляет собой цепь окислительно-восстановительных реакций с поглощением немалого количества солнечной энергии. Суммарно уравнение фотосинтеза имеет вид 6 CO2 + 6 H2O Механизм этого процесса следующий: при фотосинтезе происходит разложение воды, и образующийся при этом водород идёт на восстановление углекислого газа. Происходит превращение кинетической энергии солнца в потенциальную химическую энергию, которая аккумулируется сложными органическими веществами. Фотосинтез имеет огромное биологическое значение: благодаря ему ежегодно связывается около (15÷20)·1010 тонн углерода и выделяется в атмосферу 4·1011 тонн кислорода. Фотосинтез проходит в хлоропластах – клеточных органоидах, главной составной частью которых является хлорофилл. Главным продуктом фотосинтеза являются углеводы (Гексозы). Содержание минеральных элементов в растении. Их физиологическая роль, накопление в отдельных органах, потребление в ходе развития растений. Физиологические основы применения удобрений. МИНЕРАЛЬНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ — химические элементы, усваиваемые растениями. Обнаруживаются в золе при их сжигании. В корнях и стеблях травянистых растений минеральных веществ содержится до 4 — 5%, в листьях — 10 — 15, в семенах — до 3, в коре древесных растений — до 7, в древесине — около 1%. Содержание минеральных веществ в растениях может резко меняться в зависимости от состава почвы, др. условий и особенно от условий влажности. М. э. р. по их содержанию делят на макроэлементы, микроэлементы и ультрамикроэлементы. Макроэлементы — химические элементы, усваиваемые растительными клетками в больших количествах, содержание их выражается величинами от десятков до сотых долей % (Fe, Ca, К, Mg, Na, P, S, Al, Si, Cl). К микроэлементам относится любой хим. элемент, содержащийся в среде обитания и в растениях в количествах менее 0,1%, но необходимый для нормальной жизнедеятельности. Микроэлементами следует называть химические элементы, находящиеся в растениях в конц. от 10-2 до 10-5% (Мn, В, Sr, Cu, Li, Ba, Вr, Ni и др.). Ультрамикроэлементы накапливаются в клетках в концентрациях менее 10-6% (As, Mo, Co, I, Pb, Hn, Ag, Au, Ra и др.). Некоторые растения способны концентрировать определенные минеральные элементы: например, морские водоросли — бром, йод, кукуруза — золото, астрагалы — селен, сфагнум — серебро, представители семейства вересковых и брусничных — марганец. Минеральные элементы в растениях способны образовывать комплексы с веществами орг. природы. Это характерно для железа и таких микроэлементов и ультрамикроэлементов, как медь, кобальт, цинк, марганец, и некоторых др. Они входят в состав или активируют до 300 ферментов. Например, медь — компонент полифенолоксидазы, железо — пероксидазы и каталазы, кобальт — витамина В12.Известны металлорганические соединения и неферментативного характера, но с высокой биологической активностью. Это, например, соединения металлов с нуклеиновыми кислотами, белками, флавоноидами, магний входит в состав хлорофилла, медь — в состав купропротеина, кремний включен в кремнегалактозный комплекс, бор — в комплексы с флавоноидами и др. Высокая биологическая активность минеральных элементов проявляется, вероятно, и при использовании некоторых лекарственных растений, морских (бурых) водорослей при лечении тиреотоксикоза; сфагнума при лечении ран; зайцегуба опьяняющего, богатого кальцием, при лечении внутренних кровотечений; листьев черники, богатых марганцем, при лечении первых стадий диабета, для которого характерно падение содержания марганца в крови, и т. д. Поступление веществ в корневую систему: основные этапы, затрата энергии, пути и механизмы продвижения веществ до сосудов ксилемы. Зависимость поступления веществ в корень от внешних и внутренних факторов. Корни поглощают вещества из почвенного раствора (водная фаза) и при контакте с частицами ППК — почвенного поглощающего комплекса (твердая фаза почвы). ППК — это мелкодисперсная коллоидная часть почвы, смесь минеральных (алюмосиликатных) и органических (гуминовых) соединений. Большая часть коллоидов почвы заряжена отрицательно, на их поверхности в адсорбированном (поглощенном) состоянии находятся катионы. Некоторая часть коллоидов почвы в определенных условиях может быть заряжена положительно, поэтому на них в поглощенном адсорбированном состоянии будут находиться анионы. Обменные катионы и анионы — один из важнейших источников питания для растений. Катионы и анионы, находящиеся в поглощенном состоянии на частицах почвенного поглощающего комплекса, могут обмениваться на ионы, адсорбированные на поверхности клеток корня. Так может осуществляться поступление катионов К+, Са2+, Na+ в обмен на протоны, а также анионов N03-, Р043- и других в обмен на НС03- или анионы органических кислот. Особенно эффективно идет поглощение при контактном обмене, при котором происходит обмен ионами без перехода их в раствор. Поглощенные ионы адсорбируются на поверхности клеточных оболочек ризодермы. Из адсорбированного состояния ионы могут по коре корня передвигаться двумя путями: по апопласту и симпласту. При поступлении в симпласт ионы проникают через мембрану и далее передвигаются по плазмодесмам к сосудам ксилемы. Однако такие большие молекулы как белки не могут преодолеть пространство в плазмодесмах, поэтому имеются специальные механизмы. Поступление ионов через мембрану происходит с помощью переносчиков как пассивно, по градиенту электрохимического потенциала, так и активно, с использованием метаболической энергии. Механизм этого процесса и роль транспортных белков подробно разбирались выше. Ток воды с растворенными веществами, движущийся по свободному пространству (апопласту), как бы омывает все клетки коры. На всем этом пути могут наблюдаться адсорбция веществ клеточными стенками, поступление ионов в клетки через соответствующие мембраны и включение их в обмен веществ, т. е. метаболизация. Необходимо учесть, что на пути движения по апопласту имеется преграда в виде клеток эндодермы с поясками Каспари. Передвижение через клетки эндодермы возможно, по-видимому, только через цитоплазму. Даже если признать наличие в стенках клеток эндодермы промежутков для свободной диффузии, то они настолько малы, что вещества не могут через них проникнуть. В связи с этим перенос ионов через мембраны клеток эндодермы необходим и также осуществляется с помощью переносчиков. В целом между апопластом и симпластом в корневой системе происходит непрерывное взаимодействие и обмен питательными солями. По мнению некоторых исследователей, основным для транспорта ионов является симпластный путь, отличающийся от апопластного большей эффективностью и возможностью осуществлять метаболическую регуляцию. В осуществлении этого большая роль принадлежит осморегулирующей функции вакуолей. Вопрос, что является движущей силой, заставляющей клетки корня секретировать ионы в сосуды ксилемы, является спорным. Имеется мнение, что клетки, расположенные около сосудов ксилемы, обладают более низкой метаболической активностью по сравнению с более удаленными благодаря меньшему содержанию в них кислорода. В силу низкой интенсивности дыхания эти клетки не удерживают соли и отдают их в сосуды ксилемы. Затем соли вместе с водой благодаря градиенту водного потенциала поднимаются по сосудам вверх (массовый ток). Последние биохимические исследования показывают, что важную роль в переносе ионов в ксилему имеют паренхимные клетки ксилемы. Плазмалемма этих клеток содержит протонные помпы, водные каналы и различные ионные каналы, которые специализированы в поступлении и отдаче веществ. Так, в паренхиме идентифицированы два типа ионных каналов: К-специфичные и неспецифичные. Эти каналы регулируются благодаря мембранному потенциалу и концентрации калия. Из этих данных следует, что транспорт ионов из паренхимы в сосуды ксилемы находится под контролем обмена веществ путем регуляции Н+-АТФазы плазмалеммы и ионных каналов. Азотный обмен растений Высшие растения поглощают соединения азота из почвы, в виде нитратов и аммиака. Корневая система растений хорошо усваивает нитраты, которые после ферментативного восстановления до нитритов превращаются в аммиак. Восстановление идет через ряд этапов при участии фермента нитратредуктазы. NO3- NO2- NH2OH → NH3 нитрат нитрит гидроксиламин аммиак Для восстановления нитратов необходимо присутствие донора водорода и электронов, которыми являются восстановленные никотинамиды (НАДФН2 или НАДН2), поставщиком этих соединений является процесс дыхания. Большое влияние на восстановление нитратов оказывает свет, так как используются продукты, образующиеся в процессе нециклического фотофосфорелирования (НАДФН2 и АТФ), процесс стимулируется при освещении синим светом. Восстановление нитратов у растений может осуществляться и в листьях, и в корнях, однако относительная доля участия этих органов в редукции нитратов у растений разных видов сильно варьирует. По этому признаку растения подразделяют на три основные группы: 1. Растения, практически полностью восстанавливающие нитраты в корнях и транспортирующие азот к листьям в органической форме (черника, клюква). 2. Растения, практически не проявляющие нитратредуктазной активности в корнях и ассимилирующие нитраты в листьях (дурнишник, хлопчатник, свекла, марь). 3. Растения, способные поддерживать активность нитратредуктазы и в листьях, и в корнях. Это наиболее многочисленная группа, к которой относится большинство травянистых растений, в том числе злаковые, бобовые, многие технические и сельскохозяйственные культуры. Ассимиляция нитратов в листьях на свету тесно связана с процессом фотосинтеза. Реакции фотосинтеза используются как источник АТФ для синтеза нитрат- и нитритредуктазы и транспорта нитратов, а также как источник восстановителей и субстрата для связывания конечного продукта восстановления – аммиака. Аммиак также может служить источником азотного питания для растений, при этом он поступает в растения даже быстрее чем нитраты. Накопление аммиака в клетках приводит к нежелательным последствиям, растения обладают способностью обезвреживать аммиак, присоединяя его к органическим кислотам с образованием амидов (глутамина и аспарагина). Это позволяет разделить растения на амидные, образующие аспарагин и глутамин, и аммиачные, образующие соли аммония. Образование амидов в растении начинается в процессе дыхания, где в качестве промежуточных продуктов образуются органические кислоты α-кетоглутаровая и щавелевоуксусная. Эти кислоты в результате прямого восстановительного аминирования присоединяют аммиак. HOOC∙CH2CH2∙CO∙COOH + NH3 + HАДН2 ↔ HOOC∙CH2∙CH2CH∙NH2COOH + H2O + НАД α-кетоглутаровая кислота глутаминовая кислота HOOC∙CH2∙CO∙COOH + NH3 + HАДН2 ↔ HOOC∙CH2∙CH∙NH2COOH + H2O + НАД щавелевоуксусная кислота аспарагиновая кислота Глутаминовая и аспарагиновая кислоты, присоединяя еще одну молекулу аммиака, дают амиды – глутамин и аспарагин. В реакциях образования амидов необходима энергия АТФ и присутствие ионом магния, для активации сентетаз.
Роль амидов в растении разнообразна. Это не только форма обезвреживания аммиака, это и транспортная форма азотистых соединений, обеспечивающих отток их из одного органа в другие. Амиды являются материалом для построения многих других аминокислот в процессах переаминирования. Синтез белка. Для нормального синтеза белка в растительном организме нужны следующие условия: 1) обеспеченность азотом; 2) обеспеченность углеводами; 3) высокая интенсивность процессов дыхания и фосфорелирования; 4) присутствие нуклеиновых кислот: ДНК и РНК; 5) рибосомы; 6) белки-ферменты катализаторы синтеза белка; 7)ряд минеральных элементов (магний, кальций). Образованием белка заканчивается прогрессивная ветвь азотистого обмена в растениях по схеме Прянишникова. Органические кислоты → аминирование → аминокислоты (аспарагиновая, глутаниновая, α-кетоглутаровая, щавелевоуксусная) + аммиак → глутамин и аспарагин → аминокислоты → белки. Вторая половина схемы показывает последовательность в процессе распада белков (регрессивная ветвь азотистого обмена). Белки распадаются до аминокислот, далее до аммиака, он вновь обезвреживается в виде амидов (аспарагин и глутамин). На основе этих соединений образуются аминокислоты, которые идут на построение новых белков Белки → аминокислоты → аммиак → аспарагин и глутамин → аминокислоты →белки.
Рисунок 1 – Схема превращения азотистых веществ Фитогормоны: ауксины, гиббереллины, цитокинины, ингибиторы. Механизм их действия. Образование и передвижение по растению. Применение в растениеводстве. Фитогормоны - это вещества, вырабатывающиеся в процессе обмена веществ и оказывающие в ничтожных количествах влияние на физиологические процессы. Ауксины - это вещества индольной природы. Основным фитогормоном типа ауксина является β-индолилуксусная кислота (ИУК) она синтезируется из аминокислоты триптофана. Образование идет в меристематических тканях. Передвигаются из верхушки побега вниз к его основанию, а далее от основания корня к его окончанию. Передвижение идет по проводящим пучкам. Транспорт ИУК происходит как пассивно, так и активно.После окончания реакции гормон должен быть разрушен или инактивирован (фермент ИУК-оксидаза). Кроме того, ИУК может переходить в неактивное состояние образуя комплексы с сахарами, аминокислотами и другими веще-ствами. Такие комплексы могут распадаться, и тогда активности гормона быстро восстанавливается. С помощью комплексов в органе образуется запас гормона. Механизм действия. 1)ИУК стимулирует растяжение клеток. Механизм заключается в активации протонных пом, закислении апопласта, снижении прочности клеточной стенки, поступлении воды и растягивании. Вызванное ауксином увеличение поступления воды приводит к быстрому растяжению клетки, уменьшению вязкости цитоплазмы и изменению скорости ее движения, что влияет на скорость химических реакций. 2) стимулирует синтез ДНК, РНК, белка. 3) стимулирует образование придаточных корней. 4) влияет на энергетический обмен. 5) стимулирует прорастание семян. 6)Под влиянием ауксина активируются кальциевые каналы что способствует транспорту воды и питательных веществ к месту действия гормона. ИУК: клетки и ткани, обогащенные ауксином, становятся как бы центрами притяжения этих веществ. Это приводит к усиленному росту органов. Таким образом, ауксины определяют направление транспорта веществ в растении. Регулируя распределение и транспорт веществ, ауксины определяют полярность растения. 7)Асимметричное распределение ИУК вызывает неравномерный рост стеблей, корней, результатом которого являются движения органов растения. Гиббереллины - это большая группа соединений, чтобы различать их, им присвоили определенные номера: ГК1, ГК2, ГК3 и т. д. Наиболее распространен гиббереллин ГК3 - гибберелловая кислота С19Н22О6. По химической природе гиббереллины - это тетрациклические дитерпеноиды, состоящие из четырех остатков изопрена. Синтез происходит путем превращения мевалоновой кислоты. Место синтеза - молодые листья, а в клетке - пластиды. Транспорт в растении не является полярным. Гиббереллины свободно передвигаются из листьев вверх и вниз. по ситовидным трубкам, паренхимным клеткам, по сосудам транспортируются в связанной форме - гиббереллин-глюкозиды. Инактивация гиббереллинов происходит в результате их превращения в гиббереллин-глюкозиды и сложные эфиры. Разрушаются с помощью ферментов. Мех-м действя. Наиболее типичное действие гиббереллинов - это стимуляция роста стебля Рост стеблей вызван только растяжением клеток. Гиббереллин действует и на эмбриональную фазу роста клетки, а также стимулирует рост столонов и задерживает образование клубней. Гиббереллины накапливаются в семенах, луковицах, почках, клубнях. Они активизируют гидролитические ферменты и их синтез в зерновках злаков, что вызывает более быстрые превращения запасных веществ и прорастание семян. Гиббереллины – стимулируют цветение ряда растений. Цитокинины - производные аденина, у которого аминогруппа в шестом положении замещена различными радикалами. Синтезируются из мевалоновой кислоты. Распространяется цитокинин вверх с ксилемным током (пассивно и неполярно). Синтез цитокининов происходит в разных участках растения в зависимости от этапа онтогенеза. Концентрации этих гормонов происходит благодаря соединению цитокининов с глюкозой. Мех-м действия. Главные функции цитокининов стимуляция клеточного деления (усиливают синтез белка и нуклеиновых кислот, активируют рост клеток) и дифференцировки, а также задержка процессов старения. Обладают огромной аттрагирующей способностью, т. е. притягивают ассимиляты (аминокислоты, углеводы) и регуляторные вещества к клеткам и тканям, содержащим большое количество этого гормона. Задерживают старение листьев вызывая образование новых тилакоидов гран и стромы в хлоропластах, разрушившихся при старении, усиливает синтез хлорофилла. Нарушают покой, например свежеубранных клубней картофеля, активируют прорастание семян, активируют открывание устьиц, В культуре ткани стимулируют деления клеток. Подавляют рост боковых корней. Снимают апикальное доминирование, вызванное ауксинами. Способствуют росту бессемянных плодов. К ингибиторам относят абсцизовую кислоту (АБК), этилен, брассиностероиды и др. АБК содержится в корнях, стеблях, почках, листьях, плодах, во флоэмном и ксилемном соке, но особенно много ее в покоящихся почках, семенах, клубнях. Синтезируется из мевалоновой кислоты, Транспортируется АБК по сосудам и ситовидным трубкам вверх и низ во все органы. Физиологическая роль. АБК тормозит все процессы роста: задер-живает растяжение и деление клеток у молодых проростков и в культуре ткани; ингибирует распускание почек. Вызывает покой у семян. АБК играет роль антитранспиранта(закрывает устица). АБК также способствует запасанию гидратной воды в клетке. Этилен. Он хорошо растворим в воде, поэтому может транспортироваться в водном растворе. Образуется в плодах, цветках, листьях, облиственных стеблях, корнях и семенах в наибольшем количестве этилен образуется в стареющих или созревающих тканях. Синтезируется из метионина. Этилен легко окисляется. Физиологическая роль. Этилен в норме абсолютно необходим для созревания плодов, синтезируясь в клетках самого плода. Для начала созревания его концентрация должна увеличиться в три раза. Этилен - гормон механического стресса. Этилен стимулирует увядание цветков, опадение листьев и плодов, так как он ингибирует синтез и передвижение ауксина, подавляющего опадение и вызывает образование отделительного слоя в основании черешка или цветоножки, т.е. этилен регулирует явление листопада. Способствует заживлению ран (под действием этилена активизируется особая ткань - раневая перидерма.). Тормозит рост главного и стимулирует образование боковых корней. Брассиностероиды образуются в незрелых семенах. Подавляет образование придаточных корней. Повышают устойчивость растений к низким и высоким температурам, дефициту влаги в почве и воздухе. Применение фитогор
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-20; просмотров: 82; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.146.255.127 (0.069 с.) |

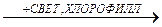 С6H12O6 + 6 O2
С6H12O6 + 6 O2