Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 8. Крупномеры страны чудес
Как и почему микроорганизмы превратились в организованные макроскопические организмы – животных и растения, столь знакомые нам по нашей повседневной жизни? На первый взгляд для такой эволюционной трансформации необходимы огромные затраты. У животных и растений размножение происходит гораздо медленнее, метаболический репертуар гораздо более ограничен, и они гораздо хуже адаптируются к изменениям внешних условий, нежели микроорганизмы. Тем не менее эти очевидные недостатки не стали препятствием на пути эволюции крупных, многоклеточных организмов. Давайте рассмотрим эволюцию сложных, или «высших», организмов и то, как они были составлены из более мелких строительных деталей, возникших тремя миллиардами лет раньше у микроорганизмов. Датировка возникновения животных и растений опирается на две независимые линии доказательств. Первая – это органические останки. Останки одноклеточных эукариотических организмов, называемых акритархами (это название в переводе с греческого означает «сомнительного происхождения»), получили относительное распространение между приблизительно 1,8 и 1,5 млрд лет тому назад. У них имелась клеточная стенка, состоящая из молекул, аналогичных молекулам целлюлозы, а также шипы и другие внешние черты, напоминающие покоящиеся споры некоторых ныне существующих одноклеточных эукариотов, таких как динофлагелляты. Хотя некоторые из акритарх, возможно, и образовывали многоклеточные колонии, до значительно более позднего времени не существует явных свидетельств существования действительно многоклеточных животных или растений.
Рис. 28. Ископаемый акритарх (Tappania plana). Этот и другие ныне вымершие организмы были предшественниками современного эукариотического фитопланктона. Этот экземпляр был найден в Северной Австралии и датируется периодом 1,4–1,5 млрд лет тому назад. Он был довольно крупным: диаметр данной клетки составляет приблизительно 110 микрометров. (Публикуется с разрешения Эндрю Нолла.)
Появление многоклеточных животных в геологической летописи выглядит так, словно они возникли ниоткуда. Дарвин понимал, что появление многих органических останков в глубочайших (а следовательно, и древнейших) из известных тогда геологических отложений в Уэльсе – в кембрийских слоях – выглядит сомнительно с эволюционной точки зрения, но он понятия не имел, как это можно объяснить.
В 1868 году шотландский геолог Александр Мюррей открыл на острове Ньюфаундленд новые ископаемые останки, залегавшие ниже кембрийских слоев. Ископаемые организмы были явно многоклеточными, но Мюррей не представлял себе, что это могло быть. Многие палеонтологи отвергли находку, считая ее артефактом. Лишь в 1957 году ископаемые, найденные в Эдиакарских холмах в Южной Австралии, были признаны как доказательство существования животной жизни в докембрийский период. Ископаемые этого периода, названные эдиакарской фауной, впоследствии были найдены в нескольких других местах земного шара, включая Белое море в России и Мистейкен‑Пойнт на Ньюфаундленде – в той самой области, что была описана Мюрреем столетием раньше. Древнейшие останки животных датируются возрастом приблизительно в 580 млн лет. Судя по всему, эти животные возникли после последнего глобального оледенения («Земля‑снежок»). Вся сохранившаяся эдиакарская фауна имела морское происхождение; это были мягкотелые организмы – они не образовывали раковин, скелетов, каких‑либо биоминералов или твердых частей, которые мы могли бы различить. Они существовали на протяжении около 90 млн лет. Эдиакарский период (эдиакарий) закончился 543 млн лет тому назад и был первой из наблюдавшихся эпох массового вымирания животных в геологической летописи. В 1909 году американский геолог Чарльз Уолкотт из Смитсоновского института случайно обнаружил в Скалистых горах на юго‑востоке канадского штата Британская Колумбия крупную свиту морских ископаемых. В целом ему удалось собрать в этом районе около 65 тысяч образцов ископаемых останков. Спустя более чем пятьдесят лет Гарри Уиттингтон и два его аспиранта опубликовали работу, в которой ясно показали, что этот разрез горных пород – сланцы Берджес‑Шейл – содержит останки, представляющие самые разнообразные типы строения организма, включая древнейших двустворчатых моллюсков, кольчатых червей и вымершие простейшие организмы с примитивными структурами, напоминающими позвоночник. Берджесские сланцы, сформировавшиеся около 505 млн лет тому назад, содержат необычайно разнообразный спектр ископаемых. На протяжении многих лет шли споры, был ли так называемый кембрийский взрыв – с виду стремительная кардинальная перемена телесного строения животных организмов, зафиксированная в геологической летописи, – артефактом захоронения ископаемых или же действительно являлся периодом увеличения животного разнообразия. Весьма вероятно, что некоторые представители эдиакарской фауны, которым удалось избежать вымирания 542 млн лет тому назад, послужили зачатками животной жизни в кембрийском периоде, однако сами эти виды‑основатели до сих пор не идентифицированы.
Рис. 29. Ископаемое Dickinsonia – вымершее животное, найденное в Эдиакарских холмах в Южной Австралии. Этот и другие представители эдиакарской фауны являются древнейшими ископаемыми животными; они появились около 600 млн лет назад в океанах. (Публикуется с разрешения Джири Липпса.)
Вторая линия доказательств относительно возникновения животной жизни не настолько однозначна. Она основывается на представлении о том, что может быть определена скорость мутаций в определенных генах, их отдельных частях или группах генов. Если мы будем знать скорость мутаций, то, сосчитав число мутаций, произошедших начиная с вымерших членов какой‑либо группы организмов, мы можем сделать заключение о скорости эволюции этой группы. Такие молекулярные часы можно использовать для обратной экстраполяции событий к точке происхождения организмов. Более поздние модели принимают во внимание вариации скорости мутаций, ввиду чего они потенциально более точны. При возможности молекулярные часы сверяют с ископаемыми отложениями, но чем дальше во времени мы пытаемся экстраполировать конкретную модель, тем она неизбежно становится все более ненадежной. Модели, основывающиеся на молекулярных часах, почти всегда показывают зарождение организмов в более раннее время, нежели свидетельства, опирающиеся на первое появление органических останков в геологической летописи. Используя модель молекулярных часов, сверенную с ископаемыми отложениями, группа ученых во главе с одним из лучших палеонтологов – специалистов по беспозвоночным Дугом Эрвином из Смитсоновского музея в Вашингтоне датировала возникновение животных временем около 700 млн лет тому назад, то есть началом эдиакария. Однако это не самый важный из их выводов. Более значительным является то, что Эрвин и его коллеги смогли также предоставить убедительные доказательства в поддержку скачкообразной эволюции животных. Таким образом, «кембрийский взрыв», судя по всему, был реально существовавшим периодом эволюции для многих новых животных форм. И хотя датировка возникновения животных имеет относительно жесткие рамки, эволюционные изменения, вызвавшие к жизни этот феномен, пока до конца не понятны. Размышляя над тем, почему животные вообще появились на планете, я часто приходил к очень простой гипотезе. Многоклеточное строение было стратегией, направленной на экологический успех в среде, содержащей очень скудное количество пищи. Попросту говоря, двигателем эволюционного отбора был голод. Нам трудно себе представить энергетику одноклеточного организма, живущего в океане. В знаменитом прекрасном эссе, написанном в честь великого физика‑теоретика Виктора Вайскопфа, его коллега Эдвард Парселл в захватывающем маленьком очерке «Жизнь в мире малых чисел Рейнольдса» описывает, какие ощущения должны испытывать микроорганизмы, пребывая в жидкости. Оказывается, для микроскопического организма вода является относительно вязкой средой. Чтобы передвигаться в такой среде, необходимо затрачивать много энергии. Парселл проводит аналогию, говоря, что клетка человеческой спермы, плывущая в воде, ощущает среду так же, как человек, плывущий в патоке. При таких условиях мы смогли бы передвигаться всего лишь на несколько метров в неделю. Однако если бы клетки смогли работать все вместе, сообща, они бы с гораздо большей эффективностью преодолевали физические барьеры, поставленные перед ними вязкостью среды их обитания.
Для того чтобы объединиться в многоклеточный животный организм, клеткам было необходимо развить у себя четыре основных свойства. Им был необходим общий для всех источник энергии. Они должны были научиться с большой точностью прикрепляться друг к другу. Они должны были распределить между собой функции, общие для всего организма, а не заботиться лишь о своих потребностях. И наконец, им надлежало воспроизводить эту схему снова, снова и снова. Все эти четыре свойства должны были работать сообща, словно отрепетированная театральная пьеса; если бы многоклеточный организм не смог выполнить хотя бы одну из этих функций, он перестал бы существовать. Вопрос об источнике энергии был наиболее критичен. За очень немногими исключениями животным, чтобы извлекать энергию из пищи, необходим кислород. У одноклеточных эукариотов кислород добирается до производящей энергию системы, митохондрии, посредством диффузии – процесса, при котором молекулы, находящиеся в постоянном хаотическом тепловом движении, перемещаются в ту сторону, где концентрация кислорода ниже. Поскольку кислород внутри митохондрий поглощается, в этой части клетки поддерживается низкая его концентрация, так что кислород перемещается из внешней среды – которой 1,8 млрд лет назад являлся океан – внутрь клетки. Диффузии вполне хватает на то, чтобы добывать кислород для одноклеточных организмов. Однако если одиночные клетки начинают укрупняться, а концентрация кислорода не очень высока, клетка будет получать недостаточно кислорода, и ее рост будет затруднен. Эта проблема еще более обостряется, когда клетки начинают формировать колонии и пытаются превратиться в многоклеточный организм.
Предположим, что существует организм, представляющий собой сплошную плоскость (типа бумажного платка), живущий на поверхности какой‑либо среды – скалы или илистой осадочной толщи. Предположим, что, подобно сложенному платку, этот организм состоит из слоев, однако вместо тонкой бумаги эти слои состоят из респирирующих клеток, как у эдиакарских ископаемых животных. Кислород проникает в верхний слой, 90 % его поглощается клетками, составляющими этот слой, и для следующего слоя клеток остается лишь 10 %. Следующий слой поглощает 90 % этих оставшихся 10 %, и третьему слою остается уже меньше 1 %. Понятно, что клеткам, расположенным у основания, кислорода почти не достанется, и они не будут функционировать как надо.
Рис. 30. Проблема диффузии кислорода в многоклеточном организме. Без какой‑либо системы циркуляции кислород мог поступать в клетки только посредством диффузии. Если животное живет на морском дне, единственным источником кислорода является вышележащая водная толща. Кислород, достигающий первого слоя клеток, уходит на нужды респирации, и второй слой получает гораздо меньше кислорода, чем первый, и так далее. Почти не вызывает сомнений тот факт, что именно диффузия кислорода послужила одной из причин эволюционного отбора плоских организмов в начале эдиакария
Ситуацию можно исправить, если изначальная концентрация кислорода будет высокой, а клетки будут организованы в такую форму, которая позволит кислороду поступать и с других сторон, или же если клетки разовьют какую‑либо систему эффективного распределения кислорода. Все эти варианты решений в конечном счете были реализованы в процессе эволюции, но изначальное условие требовало значительного увеличения концентрации кислорода в атмосфере Земли. Погребение органического вещества в океанических донных отложениях и сопутствующий приток кислорода в земную атмосферу получили резкое ускорение с эволюцией фитопланктона. В отличие от своих предков‑прокариотов, которые почти не могли погружаться в океанические глубины, поскольку были слишком малы (вязкость воды удерживала их на поверхности), эукариотический фитопланктон погружался довольно быстро. Эволюция этих организмов и последующие гибель и погребение их в донных толщах древних океанов привели к долговременной изоляции органического вещества, вследствие чего произошел скачок концентрации кислорода в планетарной атмосфере (см. главу 5). Этот скачок произошел около 700 млн лет тому назад, приблизительно через 1,7 млрд лет после Кислородной катастрофы. Второе увеличение содержания кислорода, несомненно, сыграло решающую роль в эволюции животных.
Никто не знает наверняка, каким было содержание кислорода, когда на Земле появились животные, но самые достоверные реконструкции показывают, что на кислород приходилось от 1 до 5 % объема атмосферы. На сегодняшний день это значение достигает 21 %. Есть нечто парадоксальное в том, что гибель и погребение эукариотического фитопланктона ускорили развитие кислородной атмосферы, и это привело к эволюции многоклеточных животных организмов, которые впоследствии стали питаться этим же фитопланктоном. С увеличением концентрации кислорода одноклеточные эукариоты смогли начать объединяться в колонии, поскольку проблема диффузии уже была настолько серьезной. Однако такое объединение предполагало возможность какого‑то сцепления, некоего межклеточного «клея» – второе свойство, необходимое для эволюции многоклеточных животных. Адгезивную функцию предоставлял набор из двух типов белков – коллагенов и интегринов, которые затем получат распространение среди всех животных организмов. Эти два белка действуют как пластичные эпоксидные смолы – они скрепляют клетки вместе, а также служат цементом для многих клеточных продуктов, таких как зубы, кости и раковины. Существует много типов коллагенов, однако для всех характерно наличие трех параллельных спиралей, похожих на микроскопические шурупы. Древнейшие их формы найдены у прокариотов. Коллагены известны всем – эти высушенные белки в смеси с ароматизаторами и подсластителями продаются в качестве желатиновых десертов. Коллагены стыкуются с интегринами – белками, прикрепленными к клеточным мембранам у животных. Это не единственные скрепляющие агенты, но наиболее значимые. У животных на долю коллагенов может приходиться до 25 % всех белков, имеющихся в организме.
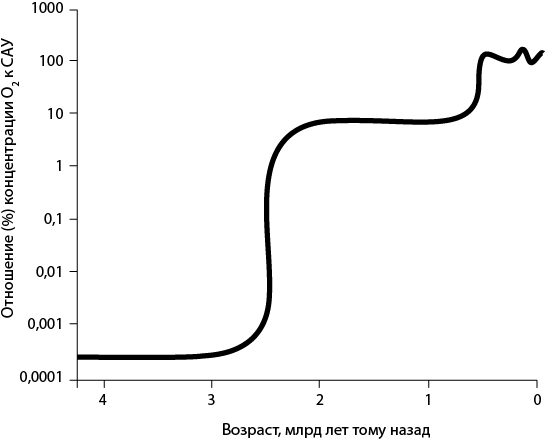
Рис. 31. Современная реконструкция содержания атмосферного кислорода на протяжении геологического времени. Обратите внимание на то, что для концентрации кислорода используется логарифмическая шкала. Концентрация кислорода на протяжении первой половины земной истории была исчезающе мала, порядка 0,0001 % современного атмосферного уровня (САУ). Во время Кислородной катастрофы (2,4 млрд лет тому назад) концентрация, вероятно, повысилась до приблизительно 1 % современного уровня и затем до примерно 10 % в эдиакарский и кембрийский периоды (600–500 млн лет тому назад). На протяжении последних 500 млн лет содержание кислорода оставалось относительно высоким и относительно стабильным, варьируя между приблизительно 50 и 150 % современного значения
И коллагены, и интегрины в той или иной форме появились в самом начале эволюции животных. Они встречаются у губок – древнейшего вида животных, где удерживают клетки в определенных положении и ориентации. По мере эволюции животного мира молекулярные клеящие средства приобретали все более важное значение, делая возможными новые, более сложные формы тела. Третье свойство – разнообразие клеточных функций – один из наиболее интересных моментов в биологии животных и растений. Даже у простейших животных и растений имеется несколько различных типов клеток. У животных существуют различные виды нервных клеток, клеток кожи, пищеварительных клеток и так далее. У растений клетки листьев, корней и побегов различаются между собой. Все эти разнообразные клетки взрослого организма происходят от одной‑единственной – оплодотворенной яйцеклетки. Независимо от того, какова функция клеток во взрослом организме, в каждой из клеток, сохранивших ядро, генетический материал идентичен тому, что содержится в остальных клетках. Вот почему мы можем взять клетки из нашей слюны, кожи, кости, печени или легких и анализировать собственный геном. Однако каждый из этих типов клеток призван выполнять свой собственный набор функций, и эти функции закодированы в генах каждого организма. Процесс, при котором клетка получает свою специализацию внутри консорции, называется дифференциацией. У животных клетки, которым еще не была определена собственная специфическая функция, называются стволовыми клетками – это клетки, которые можно убедить стать той или иной из множества типов клеток: нервной клеткой или клеткой печени и так далее. Однако откуда взялись все эти различные типы клеток в многоклеточных организмах? У цианобактерий, формирующих колонии, существуют такие клетки, которые теряют свою фотосинтезирующую способность и начинают специализироваться на связывании азота. Этот новый тип клеток крупнее, имеет более толстую клеточную стенку, а также является единственным типом клеток внутри колонии, который может связывать азот, образуя аммоний. Кроме того, такую клетку невозможно убедить снова начать фотосинтезировать – даже несмотря на то, что у нее сохранились все необходимые для этого гены. Существует несколько других примеров дифференциации. Многие одноклеточные эукариоты подвергаются тому или иному виду генетической рекомбинации и при этом трансформируют свои клетки из одной формы в другую. Генетическая рекомбинация – это такое модное словечко, означающее секс: две клетки, каждая из которых имеет половину хромосомного набора родительской клетки, комбинируют генетическую информацию, формируя новую клетку, которая начинает воспроизводиться. У одноклеточных эукариотов зародышевые клетки часто совершенно не похожи на родительские. В действительности зачатки полового размножения уходят далеко к истокам эволюционной истории; они найдены в современных эукариотических водорослях. «Споры», или зародышевые клетки, имеют половинный набор хромосом – индивидуальных сегментов генетической информации, хранящейся в ядре каждой клетки – от родительской клетки, и зачастую имеют очень различные формы. Дифференциация клеток стала фирменным знаком как животной, так и растительной эволюции. По мере развития многоклеточных организмов отдельные клетки приобретали определенные функции. У низших животных и большинства растений организм может воспроизводиться без половой рекомбинации – можно просто взять какую‑либо часть организма и вырастить ее, если иметь достаточно энергии и источников питания. В таких случаях клетки сохраняют достаточную гибкость для приобретения новых функций. Тем не менее в процессе эволюции все более и более сложных животных организмов эта гибкость была потеряна, и единственным путем возникновения новых организмов осталась половая рекомбинация – четвертое из перечисленных свойств. Половое размножение ведет к формированию оплодотворенной единичной клетки – зиготы, в которой затем происходит дифференциация в новые типы клеток по мере ее деления и развития в эмбрион. Информационная система, отвечающая за развитие и организацию клеток у животных и растений, достигла небывалой сложности, однако основной набор инструментов был позаимствован у их одноклеточных прародителей и аналогичен кворумному восприятию в микробиотических сообществах. У животных развился набор молекул, направляющий транскрипцию генов в клетках. Эти транскрипционные факторы, со временем ставшие весьма изощренными, размещают развивающийся организм животного вдоль некоторой оси и направляют деление и функционирование клеток. Так, например, у животных набор генов гомеобокса (или, на научном жаргоне, Hox‑генов) включает и выключает сотни генов во время развития эмбриона; транскрипционные факторы наподобие Hox‑генов зачастую невероятно консервативны. Впервые они были открыты в 1984 году у плодовой мушки дрозофилы, но впоследствии ученые поняли, что такие же гены встречаются во всем животном царстве – от медуз до человека. Абсолютно другой набор транскрипционных факторов развился у растений. Их представителями являются гены MADS‑бокс, организующие развитие репродуктивного аппарата. Существуют и другие, участвующие в развитии корней и побегов на ранних стадиях прорастания семян. То, что транскрипционные факторы животных и растений принадлежат к различным типам, при повсеместном распространении тех и других в соответствующих царствах, указывает на то, что молекулы, отвечающие за контроль за телесным строением этих двух групп макроскопических организмов, появились уже после того, как эти организмы пошли разными путями от последнего общего предка. Поскольку и у растений, и у животных имеются, по‑видимому, совершенно одинаковые митохондрии, маловероятно, чтобы прародителем животных был какой‑либо из протистов, потерявший пластиду. Это снова приводит нас к принуждающим факторам эволюционного отбора, которые изначально и привели к возникновению животных. Древнейшие ископаемые останки, принадлежащие к эдиакарскому периоду, не имеют очевидной связи с какой‑либо из современных форм животной жизни, однако изучение молекулярного состава показывает, что губки, сохранившиеся в геологическом разрезе начиная с кембрия, являются древнейшим из существующих типов животных. (В данном контексте под типом подразумевается просто группа животных или растений, характеризующихся сходным телесным строением. Губки принадлежат к типу Porifera; это слово означает «имеющие поры».) Строение современных губок относительно несложно. По существу эти организмы представляют собой каркас для миллионов пор, через которые протекает вода. Губка является гигантской консорцией эукариотических клеток. Их строение и стратегия питания дают ключ к вопросу о том, как и почему эти организмы возникли в эволюционном процессе. И именно здесь может оказаться информативным данное Парселлом описание жизни маленького одноклеточного организма в вязкой жидкости наподобие воды. В губках имеются клетки, по‑видимому тесно связанные с группой ныне живущих одноклеточных жгутиковых организмов, – хоанофлагелляты. У хоанофлагеллят имеется маленький воротничок, состоящий из микроворсинок, являющихся небольшими выростами клеточной мембраны. Эти организмы используют свои жгутики (от которых они и получили свое латинское название: слово flagellum означает «хлыст»), чтобы прогонять воду через эти воротнички, где микроворсинки отфильтровывают бактерии и другие мелкие органические частицы, чтобы клетка могла их поглотить. Сами жгутики представляют собой древние наномеханизмы; они найдены не только у эукариотов, но также у прокариотов, хотя строение жгутика у этих двух групп различается. У эукариотов, таких как хоанофлагелляты, жгутик состоит из девяти двойных нитей белка динеина, окружающих центральную двойную нить, состоящую из тех же молекул. Динеин – это молекулярный мотор: одна из нитей гидролизует АТФ и в этом процессе изгибается и скользит относительно соседней нити. Представим себе, как взбираемся на руках по канату (динеин): одна рука (мотор) перемещается к другой, ухватывается над ней, после чего та скользит поверх нее и ухватывается за канат, и так повторяется снова и снова. В результате жгутик хлещет взад и вперед, перегоняя воду. Жгутики такого типа появились у одноклеточных эукариотов и используются ими для передвижения в воде и питания, при котором поток частиц направляется к клетке. Этот важнейший наномеханизм впоследствии станет отвечать за множество других процессов у животных – от передвижения сперматозоидов до переваривания пищи в кишечнике. Большинство членов семейства хоанофлагеллят являются автономными одноклеточными организмами, но несколько видов могут образовывать колонии. И хотя колониальные формы одноклеточных эукариот встречаются не так уж редко, у некоторых видов хоанофлагеллят имеются гены, позволяющие им прикрепляться друг к другу с чрезвычайно высокой точностью.
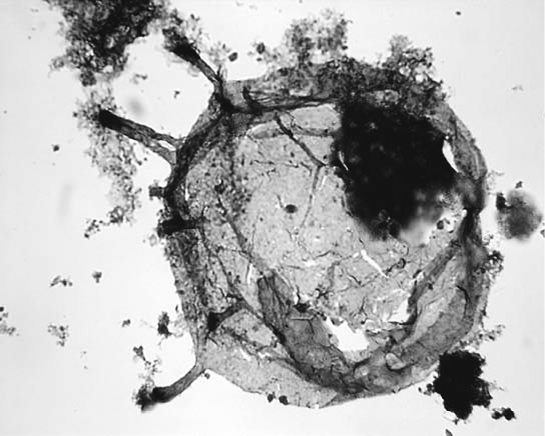
Рис. 32. Изображение колонии хоанофлагеллят (слева), где видны жгутики, при помощи которых они проталкивают бактерии и другие органические частицы внутрь воротничка для дальнейшего поглощения, а также поразительно сходные типы клеток – хоаноциты, найденные у губок (справа) В 1841 году, за девятнадцать лет до выхода в свет «Происхождения видов», французский биолог Феликс Дюжарден заметил некоторое сходство между хоанофлагеллятами и морфологией клеток, выстилающих внутренние полости губок. Он назвал эти клетки хоаноцитами. Хоаноциты губок двигают своими жгутиками в едином ритме, перекачивая через внутреннюю полость губки десятки литров воды в день. Во внутренней полости губок хоаноциты отфильтровывают из проходящей воды бактерии и органические частицы, используя свои жгутики для захвата и поглощения строительного материала для колонии. Движение жгутиков синхронизировано, чтобы создавать единонаправленный поток воды через тело животного, точно так же, как гребцы на триремах синхронизировали свои гребки, чтобы судно могло двигаться по воде. Это может показаться удивительным, но у губок нет нервной системы. Остается неясным, каким образом отдельные хоаноциты сообщаются друг с другом и какие сигналы отвечают за координацию движений миллионов жгутиков. В любом случае синхронизированное движение миллионов жгутиков способствует перемещению больших объемов воды – и в результате эта макроскопическая колония клеток более не действует так, как если бы пребывала в жидкости, вязкой как патока. Губки представляют собой микроскопический зоопарк. Поглощая от 75 до 90 % микроорганизмов, которые они фильтруют из воды, они вместе с тем дают прибежище тысячам различных микроорганизмов, с которыми находятся в отношениях взаимовыгодного симбиоза. Эти микроорганизмы обнаруживаются повсюду в миллионах мелких пор, из которых состоит тело животного. Некоторые из них обеспечивают своему хозяину пропитание, например поставляя ему витамины и другие соединения, в точности так, как поступают микроорганизмы в нашем кишечнике. Другие микроорганизмы производят токсины, защищающие животное от хищников. Собственно говоря, некоторые из наиболее токсичных молекул в животном царстве найдены именно у губок. В других случаях жильцами губки оказываются фотосинтезирующие водоросли, которые предоставляют ей источник питания, одновременно рециркулируя отходы жизнедеятельности хозяина. Сотрудничество микроорганизмов с губками стало предшественником более широких взаимовыгодных отношений, связывающих макроскопический и микроскопический миры. Эволюция губок предвещала потенциальные выгоды перехода к многоклеточному существованию. Хотя в океанах и озерах и по сей день остались хоанофлагелляты и другие эукариотические гетеротрофные организмы, координированное движение миллионов хоаноцитов позволяло губке перерабатывать гораздо больший объем воды, нежели любой одиночной клетке. По существу, даже несмотря на то что губки всю свою жизнь не сходят с одного места, благодаря ежедневной перекачке через их тела десятков литров воды их ареал добывания бактерий и других питательных частиц оказывается на несколько порядков больше, чем у их предков – плавающих одноклеточных эукариотов. Поскольку питание теперь распределялось между миллионами клеток, количество усилий, затрачиваемых на добывание пищи одной клеткой, значительно уменьшилось. Более того, благодаря тому что через организм постоянно протекал такой поток воды, снабжения кислородом оказалось достаточно для поддержания высокой скорости обмена веществ. И вдобавок ко всему, поскольку губки давали пристанище как питающим, так и токсичным микроорганизмам, они оказались более самодостаточны и защищены от хищников. Клеточное взаимодействие показало свои преимущества. Развитие строения тела животных было одним из краеугольных камней эволюции даже до эпохи Дарвина. Представление о том, что животное, создающее раковину, например двустворчатый моллюск, фундаментально отличается от животного, имеющего позвоночник, например змеи, птицы или человека, кажется очевидным на макроскопическом уровне. В этом смысле можно сказать, что мотоцикл, автомобиль, восемнадцатиколесный грузовик, океанский лайнер и реактивный самолет имеют различное строение тела, но они все имеют моторы, требующие источника энергии и использующие одно и то же топливо. Эти рукотворные машины возникли на протяжении 150‑летнего периода, и их эволюция, хотя ретроспективно она и кажется невероятно быстрой, основана на тех же основных принципах: использование однотипных механизмов для приведения в движение транспортных средств различной формы. Тот же самый принцип верен и в отношении эволюции животных. Ключевые наномеханизмы – факторы сопряжения, фотосинтетические реакционные центры, цитохромы и переносчики электронов – отвечают за поддержание жизни у всех растений и животных и развились у микроорганизмов миллиарды лет тому назад. Весь этот аппарат перешел ко многим телесным формам, прежде всего у животных. Животные – небольшая, относительно незначительная ветвь на древе жизни; они подобны множеству версий мотоциклов, машин и грузовиков, использующих в целом одинаковые моторы для того, чтобы двигаться. Фактически метаболический аппарат у животных и растений гораздо менее разнообразен, нежели он был у их микробиотических предшественников; животные не имеют доступа ко многим видам «топлива», которыми пользовались (и до сих пор пользуются) эти микроорганизмы. Зато им удалось заполучить другие инновационные процессы, отделяющие животных от их микроскопических предков. Эти инновационные процессы имели большое значение, и, хотя едва ли так уж важно перечислять их все, я хотел бы сосредоточиться на нескольких ключевых нововведениях, позволивших животным добиться такого успеха. Среди наиболее важных процессов нужно отметить продолжительную подвижность и сенсорную систему, а также формирование нервной системы и мозга. В каждом из этих случаев у системы имелся оригинал или аналог в микробиологическом мире; животные модифицировали уже существовавшие гены, им не приходилось начинать все заново. Подвижность – одна из самых ранних инноваций в эволюции животных. Хотя губки большей частью неподвижны, их близкие родственники гребневики могут плавать. Эти маленькие животные похожи на миниатюрные прозрачные футбольные мячики, но у них есть восемь рядов клеток с очень большим количеством жгутикоподобных образований – ресничек, проходящих вдоль их внешней поверхности. Эти реснички все бьются в унисон, создавая волну вдоль внешней поверхности животного, что позволяет ему перемещаться в толще воды. В какой‑то мере строение этой двигательной системы аналогично вывернутой наизнанку губке. Такая система, унаследованная от одноклеточных организмов, не особенно эффективна, и организмы по мере увеличения их размеров отказались от нее. Тем не менее ее было вполне достаточно, чтобы преодолеть проблему вязкости в мелких масштабах, с которой сталкивались в воде все одноклеточные организмы; гребневики – наиболее крупные организмы, использующие эту систему для передвижения. С развитием стрекающих – медуз и прочих – для движения стала использоваться струя воды, выталкиваемая наружу через их ротовые отверстия. Гидродинамические свойства миниатюрного футбольного мячика не так уж хороши. Все моряки мира знают это: для передвижения сквозь толщу воды подводных лодок, представляющих собой по существу вытянутые мячи, требуются большие затраты энергии. С эволюцией билатерально‑симметричных животных, таких как черви, насекомые, рыбы, рептилии, птицы и мы сами, значительная доля клеток была преобразована в мышцы, контролируемые нервами – клетками другого типа, которые благодаря скоординированным действиям способны очень эффективно двигать своего хозяина в воде или в воздухе. Для эволюции всех этих систем понадобился ряд молекулярных моторов – эта позиция была заполнена набором белков, называемых миозинами, использующими АТФ для того, чтобы «идти» по другому белку – актину. Длительное время считалось, что гены, кодирующие миозиновые белки, существуют только у животных, особенно билатерально‑симметричных. Однако по мере того как становились известны все новые и новые генетические последовательности, выяснилось, что гребневики и медузы не только содержат миозины, но что гены этих белков произошли от одноклеточных эукариотов, в первую очередь от хоанофлагеллятов. Фактически животные переняли и вторично воспользовались генами, появившимися за сотни миллионов лет до них. Механизмы одноклеточных организмов спустя миллионы лет стали снабжать энергией животных, в миллионы раз превышающих их по массе. Та же схема была обнаружена и в отношении эволюции сенсорных систем. Многие прокариотические микроорганизмы развили у себя хемосенсорную систему, являющуюся аналогом органов вкуса и обоняния у животных. Зрение – один из классических примеров того, какие трудности пришлось преодолеть, чтобы использовать унаследованные из микромира системы для более сложных организмов. На протяжении многих лет эволюция органов зрения считалась настолько сложной, что глаза могли сформироваться не иначе как под руководством божественного провидения. По всей видимости, Дарвин тоже занимался проблемой эволюции глаз, но его размышлениям по этому вопросу препятствовал недостаток информации. В первом издании «Происхождения видов» Дарвин писал: Предположение о том, что глаз со всеми его несравненными приспособлениями для фокусировки на различные расстояния, для пропускания различного количества света, для корректировки сферической и цветовой аберрации мог быть сформирован в результате естественного отбора, признаюсь откровенно, кажется в высочайшей степени абсурдным. Тем не менее здравый смысл говорит мне, что если будет доказано, что существуют многочисленные градации от совершенного и сложного глаза до глаза весьма несовершенного и простого, притом что каждая градация окажется полезной для ее обладателя (а это, несомненно, так и есть); если, далее, глаз понемногу изменяет свое строение и эти изменения наследуются (что также несомненно); если любая вариация или модификация этого органа оказывается полезной для животного в изменяющихся условиях его жизни – тогда препятствие, мешающее нам поверить, что совершенный и сложно построенный глаз мог сформироваться путем естественного отбора, хотя и непреодолимое для нашего воображения, едва ли можно считать существующим в действительности.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 79; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.14.84.29 (0.037 с.) |