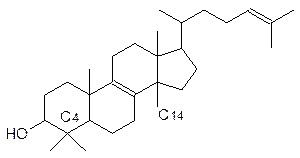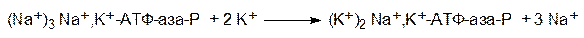Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Липидные компоненты клеточных мембран
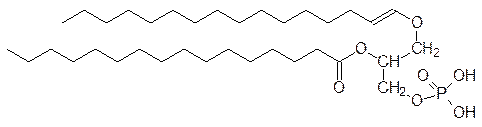
До 50% липидов клеточных мембран составляют производные фосфатидных кислот, которые представляют собой продукт этерификации диацилглицеринов фосфорной кислотой по первичной спиртовой группе, например:
В свою очередь фосфатидные кислоты можно рассматривать как продукты диацилирования L -глицерин-3-фосфата или, что то же самое, D -глицерин-1-фосфата, но стерические эффекты у жиров и производных фосфатидных кислот особой роли не играют. На основе фосфатидных кислот в организме образуется несколько важных амфифильных соединений, представляющих собой продукты этерификации одной из кислотных групп фосфорной кислоты гидроксильными группами холина, этаноламина или серина:
Эта формула соответствует фосфатидилхолину (лецитин, эфир фосфатидной кислоты и холина HOCH2CH2N+(CH3)3), другими соединениями этой группы являются фосфатидилэтаноламин (кефалин, эфир фосфатидной кислоты и этаноламина HOCH2CH2NH2) и фосфатидилсерин (эфир фосфатидной кислоты и аминокислоты серина НОСН2СН(NH2)СООН). Амфифильные свойства могут быть также обусловлены неионогеннми гидрофильными остатками. При этерификации одной из гидроксильных групп фосфатного фрагмента остатком инозита – гексагидроксициклогексана – образуется фосфатидилинозит, а его гидроксильные группы могут быть дополнительно фосфорилированы. Определенную роль в построении мембран играют и простые эфиры глицерина – вещества, построенные по типу фосфатидных производных, у которых один из остатков жирной кислоты (у первичной гидроксильной группы) заменен на енольный остаток или остаток жирного спирта (плазмалогены), например:
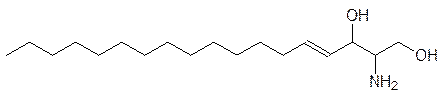
В этих веществах фрагмент фосфорной кислоты также образует эфирные связи с различными гидрофильными остатками. Близки фосфатидилхолину по биологическим функциям сфингомиелины, построенные на основе ненасыщенного аминодиола – сфингозина:
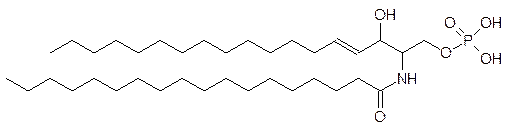
Ацилированный по аминогруппе остатком жирной кислоты сфингозин – это церамид, образующий аналогичный фосфатидной кислоте эфир с фосфорной кислотой (приведена формула фосфата церамида):
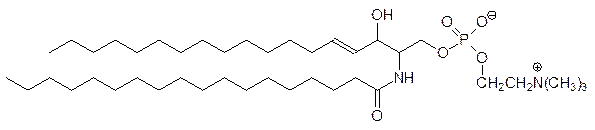
Эфир фосфата церамида и холина – это сфингомиелин:
В церамидных производных гидрофильный фрагмент может быть представлен остатками полиоксисоединений, в частности сахаров. Церамид, соединенный гликозидной связью с глюкозой – это глюкоцереброзид:
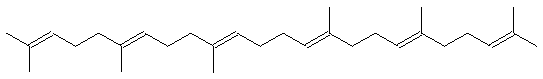
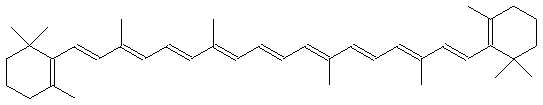
В результате присоединения к первичной гидроксильной группе церамидов олигосахаридных остатков образуются ганглиозиды, которые формируют антигенные и рецепторные участки на клеточных мембранах. В построении клеточных мембран участвуют и липиды терпеноидной природы, построенные из фрагментов изопрена. В их число входят полиненасыщенные линейные и алициклические соединения:
Сквален
β-Каротин В состав мембран входят также стероиды, например, холестерин и эргостерин:
Холестерин Эргостерин Из дегидрохолестерина (эргостерина) на свету образуется витамин D, регулирующий, в частности, кальциевый обмен. В норме в организм человека он должен поступать (или образовываться) в количестве около 10 мкг в сутки, а доза в 1,5 мг этого витамина – уже яд. Скорость образования витамина D в значительной мере зависит от светового потока, поэтому люди южных широт имеют темную кожу (светлокожие загорают), а северяне – блондины. Правда, есть еще смуглые и черноволосые эскимосы, но они получают много витамина D с рыбой и мясом морских животных. Вещества стероидной природы очень важны в составе клеточных мембран всего живого. Они определяют их лабильность, проницаемость для многих необходимых клетке веществ, для которых нет специальных транспортных систем. Если мембраны клеток животных включают преимущественно холестерин, то мембраны растительных клеток содержат большое число (в зависимости от растения) стероидов иного строения. Первичным продуктом стероидной природы, образующимся из сквалена (тритерпен линейного строения), становится чаще всего ланостерин:
Ланостерин трижды подвергается деметилированию (две метильные группы у С4 и метильная группа С14) и трансформации по боковой цепи. Окисленные стероиды с карбоксильной группой в укороченной боковой цепи – это желчные кислоты. Они играют роль поверхностно-активных веществ в пищеварительной системе. Без них скорость переваривания жиров резко снижается. Растительные клетки, наоборот, усложняют эту боковую цепь. Паразитирующие на растениях грибы чаще всего используют в качестве исходного продукта для своего мембранного стероида – эргостерина – ланостерин. Нарушение деметилирования ланостерина по С14 лежит в основе действия современных противогрибковых препаратов – фунгицидов. Стероидную природу имеют многие гормоны и токсины.
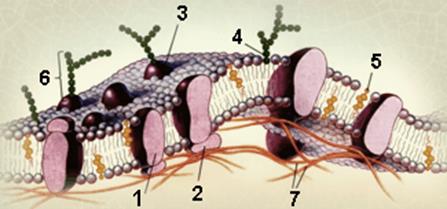
Клеточные мембраны При изучении строения клеток методом электронной микроскопии были получены экспериментальные данные, позволившие Сингеру и Николсону (1972) предложить жидкостно-мозаичную модель клеточной мембраны. На электронных микрофотографиях срезов клеток, окрашенных тетроксидом осмия и перманганатом калия, выявляются образования толщиной 6–10 нм с темными наружными (по 2–2,5 нм) и светлым внутренним (около 3 нм) слоями, которые по всем показателям соответствуют клеточным мембранам. В темных слоях повышено содержание металлического осмия. При скалывании замороженных клеток линия скола проходит по светлому слою, примерно по середине этих образований. При этом в плоскости скола обнаруживаются крупные включения, представляющие собой интегрированные в мембраны белковые молекулы. При обработке клеток растворами с высокой ионной силой от них отделяется часть связанных белков, которые удерживались на внешней стороне мембраны только ионными силами или водородными связями (периферические белки).
Представленная на рис. 3.1 мембрана состоит из бислоя липидов, в котором плавают (или закреплены) белковые молекулы, образуя в нем своеобразную мозаику. Мембранные белки могут пронизывать бислой насквозь и погружаться в него (интегральный белок 1) или примыкать к бислою (периферический белок 2). Многие мембранные белки являются гликопротеинами 3, а мембранообразующие липиды – гликолипидами 4. На рисунке показаны также холестерин 5, олигосахарид 6, элементы цитоскелета (7). Ко времени разработки этой модели было хорошо изучено строение амфифильных соединений, которые по их способности растворяться в неполярных растворителях относились к липидам, то есть жироподобным веществам. В отличие от обычных поверхностно-активных веществ выделенные из клеток амфифильные липиды имеют по два гидрофобных углеводородных остатка, которые придают им способность к образованию в воде ассоциатов с двойным слоем молекул, где полярные фрагменты обращены в сторону водной фазы, а неполярные, закрытые от воды полярными, образуют внутренний гидрофобный слой. В отличие от обычных мицелл (мицеллы образуются при ассоциации ПАВ с одним гидрофобным остатком) такие двухслойные системы могут формировать шарообразные или иные полые образования, внутри которых заключена часть захваченной водной фазы, их называют липосомами. Полученные простым смешением мембранных липидов с водой липосомы различаются по размеру в широких пределах, они могут иметь достаточно сложное строение, когда внутри одной липосомы находится несколько более мелких или когда просто идет чередование слоев с прослойками из включенной в липосому жидкости. В пределах липидного бислоя составляющие его молекулы достаточно подвижны, но перемещение одной молекулы с внешней стороны на внутреннюю и наоборот затруднено, поскольку для этого полярная часть молекулы должна пройти через неполярный слой. В общем случае мембрана представляет собой двухмерную жидкость. Углеводородные составляющие амфифильных липидов имеют разную длину, и поэтому есть взаимопроникновение «хвостов» липидных молекул одного слоя в другой, что еще более стабилизирует систему.
Включенные в клеточную мембрану белки могут перемещаться в этой двухмерной жидкости в результате латеральной диффузии. В принципе должно было бы происходить даже слипание, агрегация интегрированных в мембрану белковых молекул, но этого не происходит из-за присутствия в мембранах стероидных составляющих и фиксации белков микротрубочками и микронитями, структурирующими все внутреннее пространство клетки (это и есть цитоскелет). Мембраны разных клеток отличаются как составом входящих в их состав липидов, так и набором интегрированных и ассоциированных белков. В клетках есть также различие в составе липидов внутреннего и внешнего слоя мембраны. Это связано, во-первых, со строением и ролью белков, находящихся на внутренней и на внешней поверхности мембраны, а во вторых, с различием в составе внутренней среды клетки и окружающей ее (интерстициальной) жидкости. На внешней поверхности клетки находятся также гликолипиды и гликопротеины с олигосахаридными участками, определяющими иммунный статус клетки. Асимметрия мембран по составу липидов на примере эритроцитов такова: на внешнюю сторону мембраны, окружающей эти клетки, смотрят гидрофильные фрагменты фосфатидилхолина и ганглиозидов, с внутренней, цитозольной стороны мембраны – фрагменты фосфатидилэтаноламина (кефалина), фосфатидилсерина и сфингомиелина. Стероидные липиды служат для регуляции жидкокристаллических свойств мембран и для фиксации в них белков. Важнейшая функция мембран – это транспорт через нее веществ и частиц в обоих направлениях. Частицы, в том числе болезнетворные бактерии или вирусы, могут захватываться клеткой (эндоцитоз), когда клеточная мембрана обволакивает частицу и отделяется внутри клетки в виде пузырька (везикулы) с включением. При экзоцитозе протекает обратный процесс, при котором мембрана везикулы, образовавшейся, наример, в результате работы аппарата Гольджи, сливается с внутренней стороны с мембраной клетки и образует пору, через которую содержимое везикулы выходит в интерстициальную жидкость. Более сложен механизм обмена молекулярными веществами. Мембрана является хорошей преградой на пути диффузии полярных молекул и ионов. Плотный внутренний слой гидрофобных остатков мембранных липидов задерживает их достаточно эффективно, но диффузия ионов все же идет через небольшие поры в мембране или через участки с пронизывающими мембрану интегрированными белками. Более легко проходят через клеточную мембрану молекулы неполярных веществ, поскольку внешние гидрофильные фрагменты мембранных липидов не образуют плотного слоя. Непреодолима клеточная мембрана для любых крупных молекул. Внутренняя среда клетки – цитозоль – содержит много кислот с высокой молекулярной массой, из-за чего с внутренней стороны мембраны (если, конечно, ее целостность не нарушена) формируется отрицательный заряд, а с внешней стороны – положительный. Это является результатом пассивной диффузии катионов и небольших анионов, хотя определенный вклад в возникновение этой разницы потенциалов могут внести и белковые системы активного транспорта ионов, называемые чаще всего насосами, но общее название таких траспортных белков – транслоказы.
Переносящие ионы транслоказы предназначены для поддержания определенного водно-солевого баланса между клеткой и окружающей средой. В частности, у человека в норме концентрация ионов калия внутри клетки примерно в 40 раз выше, чем в интерстициальной жидкости, а концентрация ионов натрия в цитоплазме в 10 раз меньше, чем вне клетки. Поскольку транслоказы осуществляют активный транспорт веществ против градиента концентраций, для него нужен источник энергии. Чаще всего это энергия ангидридной связи в молекуле аденозинтрифосфата (АТФ). Так, например, Na+,K+-насос (Na+,K+-АТФаза) представляет собой комплекс белков, пронизывающих клеточную мембрану. В цитозоле специальный участок этого комплекса фосфорилируется аденозинтрифосфатом и связывается с тремя ионами натрия. Это вызывает структурную перестройку белков, в результате которой участок с ионами натрия оказывается на внешней стороне клеточной мембраны, где они обмениваются на два иона калия. Это приводит к новой структурной перестройке с перемещением участка белка с ионами калия на внутреннюю сторону мембраны. Здесь гидролитически отщепляется фосфатная группа, и ионы калия заменяются на ионы натрия. Следует новое фосфорилирование и описанная последовательность повторяется. Один цикл работы Na+,K+-АТФазы описывается следующей схемой:
Первое превращение протекает в цитозоле, второе – в интерстициальной жидкости и третье – снова в цитозоле. На поддержание водно-солевого баланса в тканях тела человека может тратиться до 40% всего образующегося в организме АТФ. Работа Na+,K+-АТФазы может быть заблокирована некоторыми соединениями стероидной природы, называемыми сердечными гликозидами. Они ингибируют гидролитическое отщепление остатка фосфорной кислоты от фосфорилированной Na+,K+-АТФазы.
|
||||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 80; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.135.200.211 (0.01 с.) |