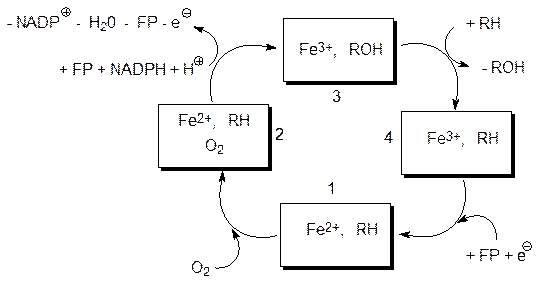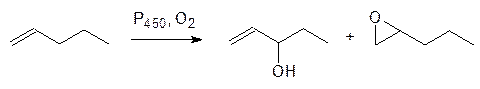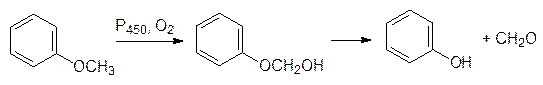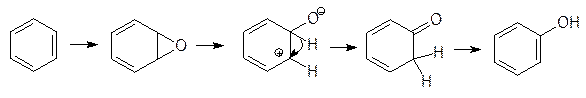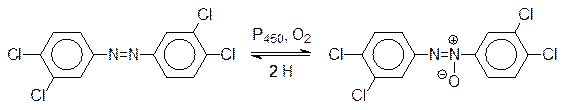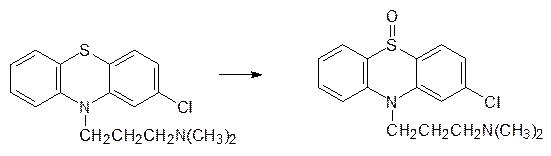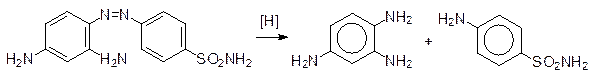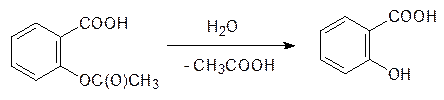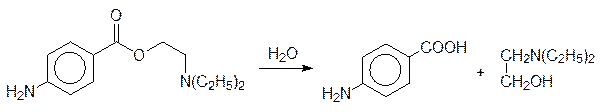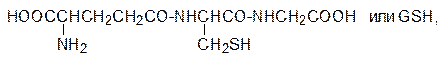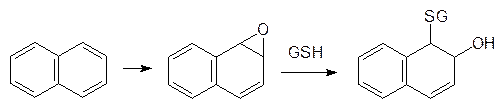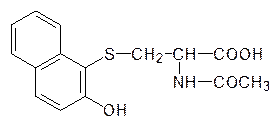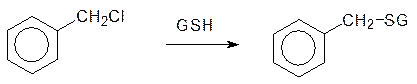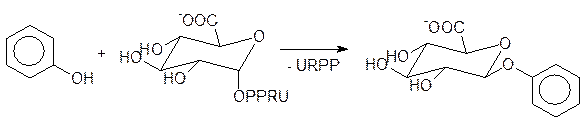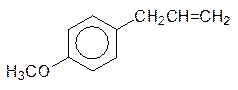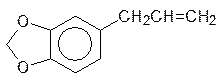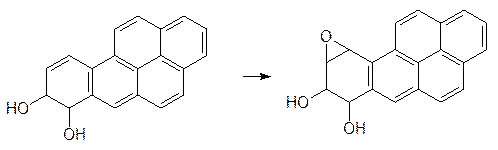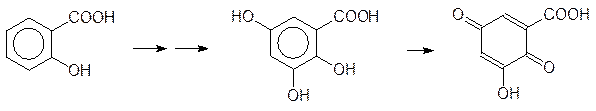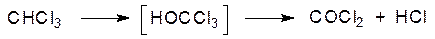Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава 12. Метаболизм ксенобиотиков
Объекты живой природы постоянно вступают в контакт с огромным числом разнообразных химических соединений. Последние входят в состав продуктов питания, содержатся в воде, поступают с вдыхаемым воздухом, и это не только необходимые организму белки, углеводы, жиры, витамины, соли, кислород, но и не представляющие пищевой ценности или даже токсичные компоненты естественного (эфирные масла, окрашенные вещества, алкалоиды, таннины, токсины и др.) или искусственного происхождения: консервирующие, вкусовые и ароматизирующие добавки, агрохимические препараты, а также оставшиеся в продуктах питания лекарственные и гормональные препараты, которые добавляли в корм птице или скоту. Независимо от их происхождения не участвующие в нормальном течении обменных процессов вещества, поступающие в организм, называются ксенобиотиками. Их отрицательное воздействие особенно сильно проявляется в индустриальных зонах с загрязненной окружающей средой. Поскольку многие ксенобиотики токсичны, живые системы для выживания должны иметь механизмы для их обезвреживания и выведения из сред, где идут нормальные метаболические процессы. У растений все отходы жизнедеятельности собираются в вакуолях – окруженных мембраной клеточных образованиях. В вакуолях действуют очень эффективные ферменты с гидролазной активностью. Нарушение целостности мембраны вакуоли приводит к гибели растительной клетки, так как содержащиеся в ней ферменты разрушают белки и другие гидролизующиеся компоненты цитозоля. Чем старше растительная клетка или чем более насыщена ксенобиотиками среда, в которой она находится, тем большее пространство клетки занимает вакуоль. У животных есть выделительная система, и ксенобиотики, так или иначе попавшие в кровь, могут быть выведены через почки. Правда, физиология почек такова, что они переводят в мочу только растворимые в воде вещества. Неполярные гидрофобные вещества образуют комплексы с транспортными белками крови и в таком виде почками не выводятся. Из этого следует, что гидрофобные ксенобиотики должны превращаться биохимическими системами в растворимые производные. Метаболизм ксенобиотиков протекает преимущественно в печени с помощью неселективных ферментов. Низкая селективность ферментов, предназначенных для метаболизма ксенобиотиков, обусловлена тем, что полный набор избирательных ферментов для всех возможных веществ (а их уже синтезировано более 10 млн.) нереализуем. Предназначенные для метаболического превращения ксенобиотиков ферменты должны быть достаточно универсальными для катализа превращений субстратов с широким набором функциональных групп в широком диапазоне молекулярных масс и пространственных особенностей. Низкая избирательность этих ферментов не позволяет им различать ксенобиотики и эндогенные вещества, и поэтому они вызывают превращение и многих необходимых для течения нормальных метаболических процессов субстратов и регуляторов метаболизма. Понятно, что в отсутствии в организме ксенобиотиков содержание этих ферментов должно быть минимальным, а появление ксенобиотиков должно индуцировать биосинтез этих ферментов.
В этом состоит первый серьезный недостаток реализуемой в живой природе системы обезвреживания ксенобиотиков – она атакует любые молекулы, в том числе и такие жизненно важные для организма вещества, как, например, гормоны, нейромедиаторы, субстраты, метаболиты, белки и многие другие участвующие в обеспечении существования организма молекулы. Но несовершенство системы, предназначенной для биотрансформации ксенобиотиков, этим не ограничивается. Введение полярных функциональных групп в инертные и неполярные молекулы чаще всего сопровождается их метаболической активацией с образованием интермедиатов с высокой реакционной способностью. Если эти промежуточные соединения не будут полностью трансформированы в течение кратчайшего времени, то появляется риск химического повреждения ими таких биомолекул, как белки, ДНК и РНК. Наибольшую опасность представляют эпоксиды, карбониевые ионы, ионы нитрония и другие электрофильные продукты превращений. Они взаимодействуют с белковыми молекулами и с нуклеиновыми кислотами, приводя к истощению иммунной системы, к нарушению нормального течения метаболизма и к мутациям в структуре ДНК. Следствием таких превращений, сопровождающих метаболизм ксенобиотиков, могут быть цирроз печени, раковые заболевания и другие повреждения систем жизнеобеспечения.
В качестве примера можно привести действие неизбирательной монооксигеназы на женский половой гормон эстрадиол. Его ароматический цикл в результате такого ферментативного окисления превращается в соответствующий ареноксид, и следствием этого становится не только снижение концентрации гормона. Гораздо более серьезные последствия связаны с высокой мутагенной и канцерогенной активностью образующегося при этом соединения:
Когда в 40-е гг. ХХ в. началось интенсивное исследование химического канцерогенеза, появилась надежда, что проблема онкологических заболеваний может быть решена в кратчайшие сроки. В основе идеи лежали представления о возможности выявления канцерогенных функциональных групп в химических соединениях. Предполагалось, что, исключив возможность контакта людей с такими веществами, можно будет избавиться от злокачественных опухолей. Но спектр веществ с канцерогенной активностью постоянно расширяется (канцерогенен даже химически инертный асбест) и, кроме того, химический канцерогенез это всего лишь один из факторов, способствующих возникновению этой болезни. На современном уровне понимания проблемы можно только утверждать, что одной из причин возникновения злокачественных опухолей является метаболическая активация клеточными ферментами поступающих в организм ксенобиотиков с образованием промежуточных продуктов, которые могут взаимодействовать с нуклеофильными центрами в ДНК. Мутагенная активность веществ может быть выявлена в достаточно простом опыте на мутантной форме бактерии Salmonella, которая нуждается для роста в гистидине. Для проверки такой штамм наносят на питательную среду, лишенную гистидина и прибавляют исследуемое вещество или исследуемое вещество вместе с гомогенизированными клетками печени крысы. Если в присутствии исследуемого вещества появляются колонии бактерий, то это означает, что прошла обратная мутация и клетки снова могут синтезировать гистидин, то есть мутагеном является само вещество. Если же такие колонии появляются только при совместном действии исследуемого вещества и гомогенизата клеток печени, то это означает, что мутагенная активность является следствием метаболической активации исследуемого вещества. Точность теста (Б. Эймс) достигает 90%. Большинство наших представлений о реакциях, сопровождающих метаболизм ксенобиотиков, является результатом изучения метаболических превращений лекарственных веществ. Эти ферментативные реакции носят общий характер для всех теплокровных, хотя, конечно, существуют и определенные количественные различия как между видами, так и между отдельными особями одного вида. На уровне отдельных индивидов важную роль в метаболизме ксенобиотиков играют генетические факторы, возраст, пол, гормональный баланс, особенности микрофлоры желудочнокишечного тракта, перенесенные болезни (особенно, если это болезни печени), образ жизни и т.д. Один из вариантов классификации путей метаболизма ксенобиотиков был предложен М. Уильямсом (1959 г.). Он выделил две фазы превращений. В первой фазе в молекулу вещества вводятся новые функциональные группы или модифицируются существующие. Во второй фазе происходит присоединение к появившимся в результате первичного превращения функциональным группам высокополярных (часто ионных) структурных элементов, например, остатков глюкуроновой кислоты, серной кислоты, аминокислотных остатков. В результате такого сочетания повышается растворимость трансформированного вещества в воде и становится возможным его выведение через почки. Подавляющее большинство поступающих в организм ксенобиотиков проходит обе фазы метаболического превращения.
Самый распространенный тип реакций первой фазы метаболизма ксенобиотиков представлен реакциями окисления. Они протекают в присутствии монооксигеназ или, иначе, оксидаз смешанных функций, называемых также цитохромами Р450 (Р450, 450 нм – это длина волны поглощаемого комплексом цитохрома и монооксида углерода света). Наряду с неизбирательными монооксигеназами, катализирующими окисление ксенобиотиков, в организме есть также избирательные ферменты этого семейства, обеспечивающие, например, превращение фенилаланина в тирозин и далее в адреналин, и участвующие в биосинтезе и в превращениях стероидов (гормоны, компоненты мембран) и т.д. Оксидазами смешанных функций эти ферменты называют потому, что в их работе участвует восстановительный потенциал в виде NADРН. Эти NADРН-зависимые ферменты катализируют большое число реакций гидроксилирования, эпоксидирования и другие превращения, используя в качестве окислителя молекулу кислорода (рис. 12.1). При этом один атом молекулы кислорода встраивается в субстрат, а второй окисляет NADРН с образованием воды. В активном центре фермента молекула кислорода связывается с атомом железа гемового кофермента и активируется для реакции с органическим субстратом. Активированный атом кислорода встраивается по связям С–Н, N–Н, S–Н, С=С и т.д. Монооксигеназа с восстановленным гемом (Fe2+) связывает в активном центре субстрат RH (1) и молекулу кислорода (2). Затем один атом молекулы кислорода при участии флавопротеинового (FP) и никотинамидадениндинуклеотидфосфатного (NADPH + H+) комплекса ферментов окисляет субстрат, второй атом кислорода образует воду (железо при этом окисляется и остается один электрон) (3). После этого окисленный субстрат (ROH) отходит от активного центра, его место занимает окисляемый субстрат (RH) (4), а оставшийся на стадии 2 ® 3 электрон восстанавливает атом железа и система возвращается в исходное состояние (1).
Обеспечивающая окислительную трансформацию ксенобиотиков ферментная система цитохрома Р450, как уже отмечалось выше, относится к индуцируемым ферментам. Это означает, что содержание фермента и его активность повышаются в ответ на появление в организме ксенобиотиков. Для этого используется обычный механизм активации генов, отвечающих за биосинтез белков. Прошедший через липидный слой мембраны гидрофобный ксенобиотик связывается с рецепторным белком, в виде такого комплекса проходит в ядро клетки и запускает процесс транскрипции на соответствующем участке хромосомы. Образовавшаяся мРНК покидает ядро и становится матрицей для синтеза белковой составляющей монооксигеназы. Индукторами монооксигеназ являются полициклические ароматические углеводороды, многие лекарственные вещества (например, фенобарбитал), входящие в состав продуктов питания флавоны, производные индола, ксантины и др. Монооксигеназы, предназначенные для биотрансформации ксенобиотиков, локализованы в гладком эндоплазматическом ретикулуме клеток печени. При гомогенизации ткани печени этот ретикулум образует микросомы (отсюда еще одно название таких монооксигеназ – микросомальные оксигеназы). В реакциях биохимического окисления алканов и алкильных остатков сложных молекул гидроксилированию подвергаются концевые группы или предпоследний атом углерода в цепи. Так, например, основным продуктом метаболического превращения гексана является гексан-2-ол. С соединениями, содержащими функциональные группы, реакция идет по функциональной группе или по ближайшему к функциональной группе атому углерода, например:
В соответствии с этим олефины окисляются в аллильное положение или эпоксидируются, а анизол окисляется по метильной группе с образованием полуацеталя, который распадается на фенол и формальдегид, то есть идет реакция О‑деалкилирования:
По аналогичной схеме жаропонижающее средство фенацетин деалкилируется с образованием парацетамола (панадола):
Ароматические углеводороды также окисляются в реакциях с участием цитохромов Р450. Бензол и многие замещенные бензолы превращаются при этом в фенолы. Сначала предполагалось, что реакция протекает просто путем внедрения атома кислорода по С–Н-связи, но затем было показано, что промежуточными продуктами этой реакции являются ареноксиды. Перегруппировка ареноксидов в фенолы или NIH‑сдвиг (NIH-Shift, National Institute of Health, США) протекает без участия ферментов:
Это было доказано элегантными опытами с дейтериевыми метками. NIH-сдвиг включает гетерогенный разрыв одной из связей эпоксида, сдвиг гидрид-иона в соседнее положение и таутомерное превращение образовавшегося ненасыщенного кетона с восстановлением ароматической структуры. Образующийся из бензола ареноксид – сильнейший алкилатор, его токсическое действие особенно ярко проявляется на клетках костного мозга, поэтому везде, где это возможно, бензол надо заменять толуолом. В отличие от бензола толуол окисляется монооксигеназами с образованием гораздо менее токсичного бензилового спирта.
В молекулах с аминогруппами окисление может идти как по атомам углерода, так и по атому азота. В первом случае идет реакция деалкилирования, а во втором образуются токсичные гидроксиламины, например:
Очевидно, что монооксигеназы могут окислять и атомы азота в пептидных связях белков с образованием токсичных и канцерогенных соединений с CO-N(OH)-группами. Третичные амины могут быть окислены до N-оксидов. Одним из серьезных экологических загрязнителей является 3,4,3¢,4¢-тетрахлоразобензол, образующийся в природе из агрохимических препаратов на основе 3,4-дихлоранилина. Этот замещенный азобензол является индуктором образования монооксигеназ, но его ферментативное окисление по пассивированным заместителями ароматическим С–Н-связям практически не идет. В организме теплокровных он превращается в N-оксид, который снова легко восстанавливается NADH-зависимыми ферментами в соответствующий азобензол:
Из-за этого его превращение в растворимую в воде форму и выведение оказывается невозможным, и он накапливается в организме, создавая при этом повышенный фон монооксигеназ, что, как уже отмечалось выше, приводит к истощению ресурса иммунной системы и ко всему спектру связанных с этим заболеваний, включая злокачественные опухоли. Тяжелая экологическая ситуация в района Арала связана с загрязнением вод 3,4,3¢,4¢-тетрахлоразобензолом. Еще более мощным индуктором биосинтеза монооксигензаз является очень токсичный 2,3,6,7‑тетрахлордибенздиоксин, образующийся из хлорфенолов и при сжигании хлорированных органических соединений:
Атомы серы в ксенобиотиках и в различных биохимических структурах легко окисляются монооксигеназами. Лекарственный препарат хлорпромазин (аминазин), используемый для лечения некоторых психических заболеваний, окисляется цитохромами до соответствующего сульфоксида:
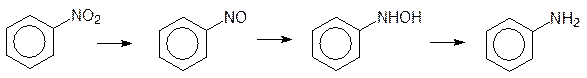
Второй тип реакций первой фазы биотрансформации ксенобиотиков – это восстановление. Реакции восстановления ксенобиотиков встречаются гораздо реже, чем реакции окисления, но этот механизм превращений важен для перевода в амины нитро- и азосоединений. Восстановление нитрогрупп катализируется нитроредуктазами и оно также идет с промежуточным образованием токсичных нитрозосоединений и гидроксиламинов.
Восстановительное расщепление азогруппы было изучено на примере азокрасителя красного цвета пронтозила («красный стрептоцид»), показавшего хорошие бактерицидные свойства. Оказалось, что сам краситель малоактивен, но в бактериях он легко восстанавливается с разрывом связей между атомами азота и превращается в триаминобензол и бактерицидный амид сульфаминовой кислоты («белый стрептоцид»), который представляет собой антиметаболит п -аминобензойной кислоты, участвующей в бактериях в биосинтезе фолиевой кислоты:
Стрептоцид – один из известнейших соединений-лидеров в синтезе большого числа антибактериальных препаратов из группы сульфамидов. Третий путь превращений в первой фазе биотрансформации ксенобиотиков представлен реакциями гидролиза. На сложных эфирах они протекают при катализе эстеразами, на амидах – амидазами. Эти ферменты присутствуют в крови, в печени и во многих тканях тела. Скорости гидролиза подчиняются общим химическим закономерностям: эфиры гидролизуются легче, чем амиды, а стерические факторы и в ферментативном гидролизе очень важны. Самый известный пример такого превращения представлен гидролизом ацетилсалициловой кислоты (аспирина) с образованием салициловой кислоты, то есть по современной фармакологической терминологии аспирин представляет собой пролекарство, так как именно салициловая кислота отвечает за лекарственную активность аспирина:
Известное местное обезболивающее средство – новокаин – гидролизуется с образованием п -аминобензойной кислоты и диэтиламиноэтанола:
Новокаин блокирует болевые ощущение на один–два часа. Другой гораздо более эффективный местный анестетик лидокаин, амид 2,6-диметиланилина и N,N‑диэтилглицина, обеспечивает блокаду болевых ощущений на время до шести часов:
Анилид диэтилглицина без метильных групп, экранирующих амидную связь, тоже проявляет свойства анестетика местного действия, но скорость его гидролиза в 100 раз выше, чем у лидокаина. Высокая скорость такого превращения исключает возможность использования его в качестве лекарственного средства, так как его действие ограничивалось бы минутами. Метаболическая детоксикация электрофильных ксенобиотиков (например алкилгалогенидов и эпоксидов) протекает с участием глютатиона. Этот трипептид – g‑глютамилцистеинилглицин
содержит нуклеофильную тиольную группу, которая легко вступает в реакции с алкилаторами и легко окисляется с образованием дисульфидной связи. Глютатион, для которого в биохимии используется обозначение GSН, содержится в тканях печени и почек в концентрации около 10 ммоль/л и он используется клетками не только для детоксикации электрофильных ксенобиотиков. Судя по всему, глютатион принимает также участие в регуляции окислительно-восстановительного потенциала в клетках организма и в нейтрализации свободных радикалов. На этом основано использование глютатиона в средствах для лечения онкологических заболеваний. Взаимодействие глютатиона с электрофильными реагентами было показано на примере метаболической трансформации нафталина. В организме нафталин окисляется с образованием соответствующего ареноксида, который реагирует с глютатионом по схеме:
Образующееся производное глютатиона гидролизуется по пептидным связям с отщеплением остатков глютаминовой кислоты и глицина, а связанный с ксенобиотиком остаток цистеина ацетилируется по аминогруппе, и из организма выводится уже производное меркаптуровой кислоты (N‑ацетилцистеина):
Галогензамещенные соединения с алкилирующей способностью обезвреживаются глютатионом с участием глютатионтрансфераз по обычной схеме с образованием тиоэфиров, например:
Ксенобиотики с функциональными группами и продукты их превращений в первой фазе метаболизма не всегда имеют достаточную растворимость для выведения их через почки. Для их гидрофилизации используется вторая фаза метаболизма, представляющая собой реакцию конъюгации (сочетания) с сахарами или с их производными или же с аминокислотами и простейшими пептидами. В редких случаях для этого используются также реакции сочетания с неорганическими кислотами, так как концентрация их солей в жидких средах организма поддерживается в очень узких рамках, а потери не так легко восполнить. Реакции конъюгации протекают с участием гидроксильных, аминных, карбоксилатных или тиольных групп, которые могли присутствовать в ксенобиотике изначально или образовались на первой фазе метаболизма. У теплокровных вторая фаза метаболизма чаще всего представлена образованием глюкуронидов – производных глюкуроновой кислоты по аномерной гидроксильной группе. Конъюгация метаболизированного на первой стадии ксенобиотика с остатком глюкуроновой кислоты протекает в результате взаимодействия соответствующей функциональной группы ксенобиотика с уридиндифосфатглюкуроновой кислотой, которая образуется из глюкозо-1-фосфата, АТФ и уридинмонофосфата с последующим окислением концевой гидроксиметильной группы до карбоксилатной. Так, например, фенол выводится из организма после его превращения в глюкуронид по реакции с уридиндифосфатглюкуроновой кислотой с одновременной защитой токсофорной гидроксильной группы (U – остаток урацила, R – остаток рибозы и РР – пирофосфатный остаток):
Эта реакция катализируется глюкуронилтрансферазой – ферментом, локализованным в клетках печени. Как уже отмечалось, образование сульфатов используется живой природой реже, чем образование глюкуронидов. И все же сульфатные производные в ограниченном объеме используются для выведения из организма спиртов, фенолов и ароматических аминов. Их образование включает три стадии: из сульфатного аниона и АТФ образуется аденозин-5¢-сульфат, который затем фосфорилируется с участием АТФ по 3¢-положению, и уже этот смешанный ангидрид (PAPS – 3¢-фосфоаденозин-5¢-фосфосульфат) переносит сульфатный остаток на субстрат. Вот так, например, в виде растворимого в воде сульфата выводится из организма активное начало панадола (А – остаток аденина):
Ацилирование – обычная реакция второй фазы трансформации ксенобиотиков, используемая для выведения из организма токсичных кислот. В этом случае ацилированию ксенобиотиками с кислотными группами подвергаются полярные аминокислоты (например, глицин или глютаминовая кислота), гораздо реже в этом превращении участвуют серин, аспарагиновая кислота, орнитин или таурин (H2NCH2CH2SO3H). В отличие от предыдущих реакций глюкуронирования или сульфатирования в этом случае активируется сам ксенобиотик. Его кислотная группа реагирует с АТФ и коферментом А с образованием ацилированного по атому серы кофермента А. Тиоэфиры являются хорошими ацилирующими средствами, тем не менее перенос ацильных остатков на соответствующие аминокислоты катализируется специфичными ацилтрансферазами. Так, например, токсичная бензойная кислота выводится из организма в виде гиппуровой кислоты, образующейся по схеме:
Как отмечалось выше, взаимодействие ксенобиотиков с монооксигеназами может приводить к продуктам, которые опаснее исходных соединений. Так, например, с образованием эпоксидов чаще всего окисляются входящие в состав душистых масел растений замещенные аллилбензолы:
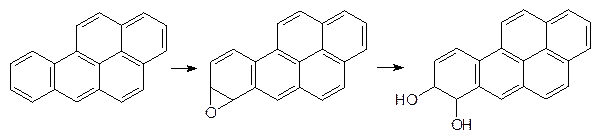
Представленные вещества, эстрагол и сафрол, содержатся в эфирных маслах укропа, сельдерея и других растений с ароматными листьями. Образующиеся из них эпоксиды обезвреживаются глютатионом, но часть их реагирует с биохимическими мишенями, образуя аллергены и вызывая мутации. Образование ареноксидов и эпоксидов лежит в основе канцерогенного действия полиядерных ароматических соединений. В продуктах пиролиза органических соединений (табачный дым, выхлопные газы) содержатся значительные количества бензопирена. Это вещество окисляется по 7,8-положению с образованием эпоксида, оксирановый цикл которого легко раскрывается эпоксидгидратазой:
Однако при повторном акте окисления образуется эпоксид, который уже не гидролизуется эпоксидгиратазами:
Он встраивается по принципу интеркаляции в ДНК и после этого алкилирует азотные функции, вызывая химические превращения этого носителя наследственной информации. До 80% поступившего в организм подопытных животных бензопирена реагирует по этой схеме. Еще один пример токсического превращения в присутствии монооксигеназ показан на парацетамоле. Этот широко рекламируемый мягкий анальгетик можно купить без рецепта, но в высоких дозах парацетамол вызывает поражения тканей печени и почек. Известны даже случаи летальных исходов от передозировки этого лекарства (вкусный детский панадол!). Как было показано выше (см. стр. 242), это вещество вступает в реакции сочетания с образованием глюкуронида или сульфата, но наряду с этим идет и реакция окисления по атому азота с образованием замещенной ацетгидроксамовой кислоты, которая дегидратируется и превращается в токсичный N‑ацетилхинонимин:
N-Ацетилхинонимин легко присоединяется к нуклеофильным центрам с восстановлением ароматической структуры. Понятно, что его действие на белки приводит к образованию аллергенов, а ДНК подвергается мутациям. Альтернативный анальгетик аспирин гидролизуется с образованием салициловой кислоты, которая окисляется с образованием полигидроксибензоатов. Их окисление также может приводить к токсичным хинонам, например:
Такая активация салициловой кислоты может лежать в основе так называемой аспириновой астмы, представляющей собой аллергическую реакцию на аспирин. Еще один вариант окислительных превращений связан с галогенированными алифатическими соединениями. Известна высокая гепатотоксичность многих хлорированных углеводородов. Полного понимания механизма этого эффекта пока нет, но можно предположить, что в его основе лежит образование веществ с ацилирующей способностью и свободных радикалов. В простейшем случае это можно представить на примере окисления хлороформа, который может превращаться в фосген:
В заключение этого раздела можно сказать, что точного представления обо всех механизмах биохимической трансформации ксенобиотиков пока получить не удалось. И все же приведенные здесь примеры позволяют оценить риск, связанный с работами по синтезу новых химических соединений, и понять необходимость продуманного подхода к проблемам практического использования веществ, которые могут подвергаться биохимически превращениям с образованием токсичных веществ или индуцировать образование неизбирательных монооксигеназ.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 164; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.220.16.184 (0.041 с.) |