
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Источники ДНК для клонирования
Существуют три источника молекул ДНК, исполь- зуемых в генетической инженерии: фрагменты генети- ческого материала различных организмов; двунитевые ДНК, полученные на основе однонитевой ДНК, ком- плементарной мРНК (кДНК); ДНК, полученная путем химико-ферментативного синтеза. Получение кДНК необходимо для экспрессии в бактериях генов белков человека: гены эукариот содержат интроны, а в клет- ках бактерий отсутствует механизм, обеспечивающий сплайсинг, поэтому применяют специальные приемы для получения ДНК, состоящей из последовательно- стей нуклеотидов, соответствующих экзонам. Для синтеза кДНК в качестве матрицы использу- ют зрелую мРНК (не содержащую интронов), которую предварительно полиаденилируют (рис. 63). Далее ее отжигают с олиго- dT, который служит затравкой при копировании нити обратной транскриптазой. От- Рекомбинантные белки − коммерческие и проходящие клинические испытания препараты
Таблица 20. Белок Система экспрессии Показания к применению
Инсулин E. coli Диабет Соматотропин E. coli Гипофизарная карликовость, остеопороз Интерферон α2 E. coli Лейкемия, профилактика простудных заболеваний Интерферон γ E. coli Опухолевые и вирусные заболевания Активатор тканевого плазминогена E. coli, дрожжи, клетки животных Тромбоз α1-антитрипсин E. coli, дрожжи Эмфазема Интерлейкин-2 E. coli, дрожжи, клетки животных Опухолевые заболевания Фактор некроза опухоли E. coli, клетки животных Опухолевые заболевания Сывороточный альбумин человека дрожжи Плазмозаместительная терапия
Фактор VIII клетки животных Гемофилия Фактор IX то же Болезнь Кристмаса
ДНК плазмы сайт EcoRI
расщепление рестриктазой EcoRI расщепление рестриктазой EcoRI ДНК
Рис. 63. Схема синтеза двуцепочечной кДНК на мРНК.
Различают три основных класса рестриктаз. Все они узнают на двуспиральной ДНК строго опреде- ленные последовательности нуклеотидов. Однако смещение, отжиг, обработка ДНК-лигазой сайт EcoRI
комплементар- ные «липкие концы» рестриктазы класса I разрывают молекул ДНК в про- извольных точках, а классов II и III — в строго опре- деленных точках внутри сайтов узнавания или на фик- сированном от них расстоянии. Ферменты классов I Amp’
трансформация, отбор клонов, несущих гибридную ДНК
хромосомная ДНК
чужеродная ДНК сайт EcoRI
гибридные плазмиды и III имеют сложную субъединичную структуру и об- ладают двумя типами активностей — метилирующей и эндонуклеазной. Ферменты II класса состоят из двух отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы. По этим причинам в генетической инженерии используют исключитель- но рестриктазы класса II. Существует общепринятая номенклатура, в со- ответствии с которой рестриктазы и метилазы обо- Рис. 62. Схема типового опыта по генной инженерии: Ampr — устойчивость к ампицилину — генети- ческий маркер плазмиды.
Рестриктазы
Ферменты рестриктазы являются необходимым инструментом при манипулировании с генами. Это специфические эндонуклеазы, которые являются со- ставной частью системы рестрикции — модификации прокариотических клеток. Эта система связана с защи- той клеток от проникновения чужеродной ДНК. Си- стема модификации осуществляет метилирование собственной ДНК немедленно после репликации. Чу- жеродную ДНК, проникающую в клетки, бактерии ги- дролизуют с помощью рестриктаз. Эти эндонуклеазы связываются с ДНК в определенных участках (сайты узнавания) и расщепляют ее на фрагменты (рестрик- ты). Собственную ДНК рестриктазы не разрушают, поскольку ее сайты узнавания метилированы. Систе- мы рестрикции-модификации обнаружены у всех ис- следованных бактерий и у некоторых дрожжей. значаются, соответственно, буквами R и M. Название фермента складывается из первой буквы рода и двух первых букв вида бактерии, из которой он был вы- делен, например, Bacillus subtilis — Bsu, Escherichia coli — Eco. При необходимости дается типовая ха- рактеристика штамма, например, Hinc — фермент из Haemophilus influenzae, серотип C. Если в опреде- ленном штамме бактерии имеется несколько систем рестрикции, дается дополнительно цифровое обозна- чение. Если фермент закодирован в генах плазмиды или фага, к названию фермента добавляется название внехромосомного элемента: EcoRI, EcoP I — ферменты E. coli, кодируемые плазмидой RI и фагом PI, соответ- ственно. В табл. 21 приведена характеристика некоторых рестриктаз, используемых в генетической инжене- рии. Разрывы цепей ДНК могут происходить по оси симметрии, и тогда образуются фрагменты с тупыми концами (например, рестриктаза BalI), либо на некото- ром расстоянии от оси, и тогда образуются фрагменты с однонитевыми липкими 5′ (рестриктазы EcoRI) или 3′ (рестриктазы PstI) концами. Рестриктазы, используемые в генной инженерии.
Таблица 21. менты, способные соединять полностью двунитевые фрагменты, однако эта реакция протекает лишь при Обознечение рестриктазы Последова- тельность, узнаваемая рестриктазой Обозначение рестриктазы Последова- тельность, узнаваемая рестриктазой высоких концентрациях ДНК и лигазы [27]. При объединении ДНК донора и вектора при от- жиге и лигировании образуются не только гибридные
1 2 3 4 молекулы, но и исходные векторы, что затрудняет даль- EcoRI G AATTC CTTAA G Hind III A AGCTT Xho l C TCGAG GAGCT C Hind II GPyC GPuC нейшую работу по клонированию ДНК донора. Поэто- му разработаны специальные методы, позволяющие направить процесс преимущественно на получение
TTCGA A CPuG CpyG 1 2 3 4
В настоящее время выделено более 500 рестриктаз класса II, однако среди них имеются ферменты, кото- рые узнают в ДНК одни и те же последовательности. Такие группы называют изошизомерами. Различают ис- тинную изошизомерию, когда ферменты узнают одну и ту же последовательность нуклеотидов и разрывают ДНК в одних и тех же точках, и ложную, когда фермен- ты, узнавая один и тот же сайт на ДНК, производят раз- рывы в разных точках в пределах того же сайта. Если предположить, что участки распознавания расположены в цепи ДНК случайно, то мишень для ферментов, узнающих сайт из четырех нуклеотидов, должна встречаться в среднем один раз через каждые 256 по (пар оснований), а для ферментов, узнающих лишь шесть нуклеотидов — через 4096 по. Очевидно, что если сайт рестрикции окажется внутри гена, об- работка рестриктазой приведет к его инактивации. Ве- роятность такого события очень велика при использо- вании мелкощепящих рестриктаз и значительна — при употреблении крупнощепящих (узнающих шестерки нуклеотидов). Поэтому для получения неповрежден- ного гена проводят обработку поочередно нескольки- ми крупнощепящими рестриктазами, либо применяют прием недорестрикции, т. е. обработку в таких услови- ях, когда происходит расщепление лишь в одном сайте из нескольких возможных. Особую ценность представляют рестриктазы, под действием которых образуются фрагменты с самоком- плементарными липкими концами: они эффективно используются при конструировании рекомбинантных молекул.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 74; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.237.255 (0.019 с.) |



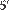
 жиг — это процесс реассоциации ДНК с образованием
жиг — это процесс реассоциации ДНК с образованием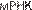 водородных связей между парами оснований. После
водородных связей между парами оснований. После 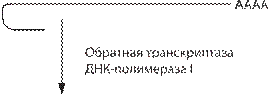
 обработки обратной транскриптазой мРНК удаляют щелочным гидролизом. Образовавшаяся кДНК име- ет на 3′-конце шпилечную структуру, которая может служить затравкой при построении второй нити ДНК с помощью ДНК-полимеразы. Для удаления олиго- dT и однонитевой петли используют S1 -нуклеазу, расще- пляющую однонитевую ДНК.
обработки обратной транскриптазой мРНК удаляют щелочным гидролизом. Образовавшаяся кДНК име- ет на 3′-конце шпилечную структуру, которая может служить затравкой при построении второй нити ДНК с помощью ДНК-полимеразы. Для удаления олиго- dT и однонитевой петли используют S1 -нуклеазу, расще- пляющую однонитевую ДНК. сайт EcoRI
сайт EcoRI
 чужеродная
чужеродная







