Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекция №11 морфо-функциональная организация ствола мозга, промежуточного мозга и мозжечка
План лекции
Структурно-функциональная организация Варолиева моста; Структурно-функциональная организация среднего мозга; Структурно-функциональная организация мозжечка; Структурно-функциональная организация промежуточного мозга; Ствол мозга - это часть головного мозга, включающая в себя продолговатый мозг, Варолиев мост и средний мозг. Тесные морфофункциональные связи со стволом мозга имеют промежуточный мозг и малый мозг или мозжечок. Все стволовые и связанные с ними образования выполняют три основные функции: рефлекторную, проводниковую, регуляторную. Основная часть структур ствола мозга теряет сегментарное строение, присущее спинному мозгу. Лишь каудальные отделы продолговатого мозга, граничащие со спинным мозгом имеют отдельные черты сегментарного строения, что нашло отражение в топографии ядер черепно-мозговых нервов. 1. Структурно-функциональная организация продолговатого мозга Продолговатый мозг - это самая каудальная часть стволовой части мозга, расположенная между мостом и спинным мозгом. В продолговатом мозгу расположены с IX по XII пары ядра черепно-мозговых нервов, разделенные проводящими путями, проходящими через продолговатый мозг, как в восходящем, так и в нисходящем направлении. Ядра IX пары (n. glossopharyngeus) - языкоглоточного нерва связаны с иннервацией мышц и слизистых оболочек ротовой полости. Принимают участие в реализации различных пищевых рефлексах: жевательном, глотательном, слюноотделительном. Ядра X пары (n. vagus) - блуждающего нерва связаны с иннервацией большинства внутренних органов (сердца, легких, желудка, кишечника и т.д.). Блуждающий нерв является самым мощным парасимпатическим нервом автономной нервной системы. Ядро XI пары (n. accessorius) - добавочного нерва является двигательным. Иннервирует скелетные мышцы - грудино-ключично-сосцевидную и трапецевидную. Участвует в реализации двигательных актов поворота и наклона головы, поднятии плечевого пояса вверх, приведение лопаток к позвоночнику. Ядро XII пары (n. hypoglossus) - подъязычного нерва является двигательным. Иннервирует язык, участвуя в регуляции его активности. Кроме того, в продолговатом мозге находятся скопления нейронов, участвующих в регуляции диаметра артерий. В связи с этим данное скопление нейронов, получило название сосудо-двигательного центра. В продолговатом мозге обнаружены нервные центры, обеспечивающие реализацию дыхательных рефлексов. При разрушении данных центров происходит остановка дыхания.
Таким образом, следует отметить, что в продолговатом мозге находятся нервные центры, которые принимают участие в нервной регуляции таких жизненно важных функций, как дыхание, кровообращение, пищеварение. Кроме того нервные центры продолговатого мозга принимают участие в регуляции двигательной активности и мышечного тонуса. Ретикулярная формация продолговатого мозга В продолговатом мозге имеется так называемая сетчатая или ретикулярная формация, описанная в конце 19 столетия в работах известного немецкого нейроанатома Дейтерса. Это образование было названо сетчатым за характерную особенность нервных клеток - большое количество отростков и контактов между клетками. Позже, в середине 20 столетия исследование морфо-функциональ-ных особенностей РФ проводилось известным нейрофизиологами американцем Гарацием Мэгуном и итальянцем Джузеппе Моруцци. Ретикулярная формация появляется на уровне продолговатого мозга, ее образования представлены в мосту, среднем и промежуточном мозге. Нейроны ретикулярной формации характеризуются определенными особенностями: 1) для нейронов ретикулярной формации характерна полимодальность, т.е. они принимают сенсорные возбуждения от нескольких рецепторных путей; 2) возбуждение через структуры ретикулярной формации распространяется с большим латентным периодом, поскольку оно проходит через большое количество синапсов; 3) нейроны ретикулярной формации на своих мембранах имеют большое количество рецепторов, чувствительных к содержанию ионов водорода, углекислого газа, различным лекарственным препаратам (барбитуратам, адренолитикам, адреномиметикам и другим). 4) Нейроны ретикулярной формации характеризуются значительной тонической активностью, т.е. способны постоянно в состоянии покоя генерировать потенциалы действия с частотой 10-15 имп/сек.
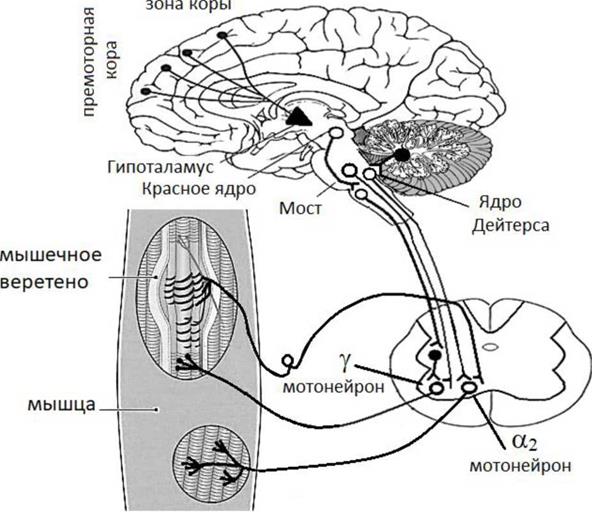
Проводниковая функция продолговатого мозга От клеток ретикулярной формации продолговатого мозга начинается латеральный ретикуло-спинальный тракт, нервные волокна которого заканчиваются на а и у мотонейронах мышц сгибателей. Ряд клеточных популяций спинного мозга получает коллатерали от спино-ретикулярного тракта, который несет информацию от температурных и болевых рецепторов. В гигантоклеточном ядре РФ продолговатого мозга заканчиваются кортикоретикулярный путь, начинающийся в сенсо-моторной коре. В этом же ядре завершается мозжечково-ретикулярный путь, связывающий ядра мозжечка и ретикулярную формацию продолговатого мозга. В продолговатом мозге заканчиваются проприоцептивные проводящие пути, идущие в задних канатиках спинного мозга (пучки Голя и Бурдаха). Кроме перечисленных выше проводящих путей через продолговатый мозг проходят транзиторно ряд проводящих путей спинного мозга начинающихся в спинном мозге и завершающихся в вышележащих образованиях (спиноталамические, спиномозжечковые, спинокортикаль-ные, спинотектальные и др.), а также ряд проводящих путей, начинающихся в вышележащих отделах ЦНС и заканчивающихся в образованиях спинного мозга (кортикоспинальные, руброспинальные, тектоспинальные, вестибулоспинальные и др.). Продолговатый мозг содержит полость - 4 желудочек, который связан с каналом спинного мозга и сильвиевым водопроводом (образование среднего мозга). Регуляторная функция продолговатого мозга Благодаря работам Р. Гранита (1955) среди образований ретикулярной формации выделяют активирующее и тормозящее ядра (вентральное и латеральное). Согласно представлениям Г. Мэгуна и Дж. Моруцци ретикулярная формация оказывает неспецифические модулирующие влияния на различные структуры центральной нервной системы. Влияния на образования спинного мозга проявляется в активации или торможении тех или иных рефлексов, в т.ч. тонических. Влияния РФ на вышележащие образования ЦНС, в частности кору больших полушарий, проявляется или в реакции десинхронизации (активации), которая характерна для бодрствующего мозга, или в реакции синхронизации (торможении), которая характерна для «спящего» мозга. В этой связи, влияя на ретикулярную формацию ствола мозга с помощью различных лекарственных средств можно регулировать тонус коры больших полушарий мозга. Один из возможных вариантов влияния нервных центров продолговатого мозга на спинальные нервные центры представлен на рисунке 11.1 отражающем влияние тормозного и активирующего ядер ретикулярной формации продолговатого мозга на у и а2 мотонейроны спинного мозга обеспечивающих мышечный тонус. моторная
Рис.3 Схема влияния нервных центров продолговатого мозга на спинальные нервные центры. (Схема регуляции мышечного тонуса по Р. Граниту)
2. Структурно-функциональная организация Варолиева моста Варолиев мост - это часть головного мозга, расположенная нейтральнее мозжечка и ростральнее продолговатого мозга. Нередко, в связи с наличием тесных морфо-функциональных отношений моста, продолговатого мозга и мозжечка эти структуры рассматривают как составные части единого образования - заднего мозга.
В Варолиевом мосту представлены ядра V - VIII пар черепно-мозговых нервов: V пара (n. trigeminus) - тройничный нерв, иннервирует жевательную мускулатуру и обеспечивает движение нижней челюсти; VI пара (n. abducens) - отводящий нерв, иннервирует мышцы глаза, участвует в управлении движением глазных яблок; VII пара (n. facialis) - лицевой нерв, иннервирует мимическую мускулатуру, принимает участие в формировании управляемых компонетов эмоций. Кроме того, в составе лицевого нерва имеются нервные волокна, относящиеся к парасимпатическому отделу автономной нервной системы, в частности иннервирующие слюнные железы; VIII пара (n. vestibulocochlearis) - предверно-улитковый нерв, связан с рецепторами вестибулярного аппарата и участвуют в регуляции позы и равновесия тела (статические и статокинетические рефлексы). Улитковые ядра связаны с рецепторами органа слуха, участвуют в реализации слухового ориентировочного рефлекса. В Варолиевом мосту имеются скопления нейронов ретикулярной формации, являющиеся продолжением ретикулярной формации продолговатого мозга. Одно из ядер ретикулярной формации моста участвует в регуляции активности а2 и у мотонейронов, иннервирующих мышечные волокна мышц разгибателей, реципрокно вытормаживая активность а2 и у мотонейронов мышц сгибателей. Таким образом, в мосту имеется ряд нервных центров, которые обеспечивают реализацию как соматических, так и автономных рефлексов. Среди соматических рефлекторных актов можно выделить и фазические и тонические рефлексы. Проводниковая функция моста аналогична проводниковой функции продолговатого мозга. 3. Структурно-функциональная организация среднего мозга Средний мозг расположен кпереди от мозжечка и Варолиева моста в виде толстостенной массы (см. рис. 2), пронизанной узким центральным каналом (сильвиевым водопроводом), соединяющим полость третьего желудочка (промежуточный мозг) с полостью четвертого желудочка
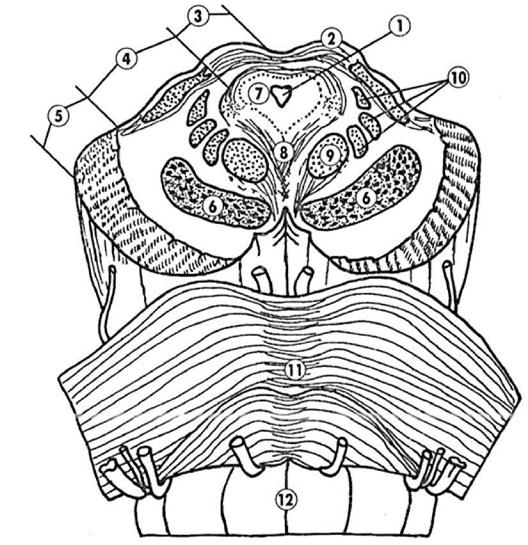
Рис. 2 Поперечный срез стволовой части мозга на уровне среднего мозга Обозначения: 1 - сильвиев водопровод; 2 -верхний холмик четверохолмия; 3-пластина четверохолмия; 4-покрышка среднего мозга; 5-основание ножки мозга; 6 - черная субстанция; 7 центральное серое вещество; 8 - ретикулярная формация среднего мозга; 9 - красное ядро; 10 - медиальная петля; 11- варолиев мост; 12- продолговатый мозг.
(продолговатый мозг). В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются латерально и образуют сетчатку обоих глаз. Сетчатка глаза, таким образом, представляет собой вынесенный на перефирию нервный центр среднего мозга. На поперечном сечении средний мозг представляет сложное образование, содержащее как скопления нейронов, так и проводящие пути. Нервные центры среднего мозга: черная субстанция, красное ядро, ретикулярная формация, четверохолмие, голубое пятно. Четверохолмие занимает верхнюю часть среднего мозга. В связи с различной ролью, выполняемой центрами четверохолмия, выделяют верхнее двухолмие и нижние двухолмие. Верхнее двухолмие играет роль зрительного подкоркового центра и служит местом переключения зрительных путей, идущих к латеральным коленчатым телам промежуточного мозга. У рыб и амфибий верхнее двухолмие достигает больших размеров и выполняет роль высшего зрительного центра. При разрушении верхнего двухолмия у рыб и амфибий возникает слепота. У человека верхнее двухолмие к тому же выполняет и роль центра ориентировочного рефлекса при действии светового раздражителя. Например, поворот головы в сторону вспышки света. Утрата верхнего двухолмия у млекопитающих не приводит к полной слепоте, но нередко приводит к нарушению ориентировочно - исследовательских реакций на незнакомые зрительные раздражители. Нижнее двухолмие выполняет функцию подкоркового слухового центра. У высших животных и человека указанные образования принимают участие в реализации ориентировочно-исследовательских реакций на действие незнакомых звуковых раздражителей. Четверохолмие и лежащие вентральнее клеточные слои среднего мозга формируют так называемую крышку среднего мозга или тектальную область. От нейронов этой области начинается так называемый текто-спинальный тракт. Вентральнее сильвиева водопровода расположены ножки мозга, которые расходясь кпереди, вступают в полушария конечного мозга. На поперечном срезе ножки среднего мозга разделяются пигментированной прослойкой (черной субстанцией) на две части: покрышку и основание ножек. Нейроны черной субстанции содержат пигмент меланин. Это филогенетически древнее образование относится к экстрапирамидной системе регуляции двигательной активности и функционально связано с лежащими в основании полушарий переднего мозга базальными ганглиями - полосатым телом и бледным шаром. В 60е годы двадцатого столетия было установлено, что в качестве медиатора нейронов черной субстанции выступает дофамин. При нарушении синтеза дофамина в нейронах черной субстанции, например, при их дегенерации, возникает тяжелое неврологическое заболевание - болезнь Паркинсона. Болезнь Паркинсона проявляется в нарушении тонких содружественных движений, функций мимической мускулатуры и в появлении непроизвольных мышечных сокращений или дрожжания (тремора). Этот синдром может быть ликвидирован при введении предшественника дофамина - L - диоксифенилаланина. В покрышке ножек среднего мозга находится парное скопление нейронов, получившее название красного ядра. Красное ядро является важным звеном переключения волокон экстрапирамидной системы, идущих от базальных ганглиев конечного мозга, а также волокон, идущих от мозжечка. Аксоны крупноклеточной части красного ядра дают начало tractus rubrospinalis, заканчивающемуся на а2 и у мотонейронах IX пластины спинного мозга. Этот тракт является конечным звеном древнейшей экстрапирамидной системы, объедняющей в единую систему регуляторные механизмы мозжечка и вестибулярные ядра. В рефлекторной деятельности среднего мозга принимают участие III пара черепно-мозговых нервов, глазодвигательный нерв (n. oculomotorius) и IV пара черепно-мозговых нервов, блоковой нерв (n. trochlearis). III, IV, и VI пары черепно-мозговых нервов медиальным продольным пучком среднего мозга объединяются в единый комплекс регуляции содружественного движения глазных яблок.
Г олубое пятно среднего мозга. Авдренэргическое образование среднего мозга. Является составной частью системы регуляции двигательной активности высших животных и человека. Считается что голубое пятно является элементом неспецифической тормозной системы мозга, имеющего связь со стриополидарной системой мозга. 4. Структурно-функциональная организация мозжечка Мозжечок или малый мозг расположен позади полушарий большого мозга, над продолговатым мозгом и мостом. Мозжечок содержит более половины всех нейронов ЦНС, хотя составляет лишь 10% головного мозга. Выделяют три основные структуры мозжечка: древний мозжечок (архиоцеребеллум), старый мозжечок (палеоцеребеллум), новый мозжечок (неоцеребеллум). Древний мозжечок включает фолликуло-нодулярную долю и нижнию часть червя. Старый мозжечок включает верхнюю часть червя, парафлоккулярный отдел, пирамиды и язычок. Новый мозжечок состоит из двух полушарий. Мозжечок не имеет прямой связи с рецепторами организма. Многочисленными путями он связан со всеми отделами центральной нервной системы. К нему направляются афферентные (чувствительные) проводящие пути, несущие импульсы от проприорецепторов мышц, сухожилий, связок, вестибулярных ядер продолговатого мозга, подкорковых ядер и коры больших полушарий. В свою очередь мозжечок посылает импульсы ко всем отделам центральной нервной системы. С различными отделами ЦНС мозжечок связан тремя парами ножек: с помощью нижних ножек мозжечок связан с продолговатым мозгом, средних - с мостом, верхних - средним мозгом. Через верхние ножки мозжечок также связан со зрительным бугром, красным ядром среднего мозга, ядрами ствола мозга, ретикулярной формацией. Через средние ножки мозжечок связан с лобными долями больших полушарий. Через нижние ножки обеспечивается связь с вестибулярными ядрами продолговатого мозга, ретикулярной формацией. Функции мозжечка исследуют путем его раздражения, частичного или полного удаления и изучения биоэлектрических явлений. Последствия удаления мозжечка и выпадения его функции итальянский физиолог Лючиани охарактеризовал знаменитой триадой А - астазия, атония и астения. Последующие исследователи добавили еще несколько симптомов - атаксию, тремор (дрожание), дисметрию (нарушение меры), у человека к тому же может наблюдаться дизартрия (нарушение речи). Безмозжечковая собака стоит на широко расставленных лапах, совершая непрерывные качательные движения (астазия). У нее нарушено правильное распределение тонуса мышц сгибателей и разгибателей (атония). Движения плохо координированы размашисты, несоразмерны, резки. При ходьбе лапы забрасываются за среднюю линию (атаксия), чего не бывает у нормальных животных. Атаксия объясняется тем, что нарушается контроль движений. Выпадает и анализ сигналов от проприорецепторов мышц и сухожилий. Собака не может попасть мордой в миску с едой. Наклон головы вниз или в сторону вызывает сильное противоположное движение. Движения очень утомляют, животное, пройдя несколько шагов, ложится и отдыхает. Этот симптом называется астенией. С течением времени двигательные расстройства у безмозжечковой собаки сглаживаются. Она самостоятельно ест, походка ее почти нормальна. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации). Как показали исследования, компенсация функций происходит за счет коры головного мозга. Если у такой собаки удалить кору, то все нарушения выявляются снова и уже никогда не компенсируются. Мозжечок участвует в. регуляции движений, делая их плавными, точными, соразмерными. Как показали исследования российского физиолога Л. А. Орбели, у безмозжечковых собак нарушаются вегетативные функции. Константы крови, сосудистый тонус, работа пищеварительного тракта и другие вегетативные функции становится очень неустойчивыми, легко сдвигаются под влиянием тех или иных причин (приём пищи, мышечная работа, изменение температуры и др.). При удалении половины мозжечка нарушения двигательных функций наступают на стороне операции. Это объясняется тем, что проводящие пути мозжечка либо не перекрещиваются вовсе, либо перекрещиваются 2 раза. 5. Структурно-функциональная организация промежуточного мозга Промежуточный мозг каудально граничит со средним мозгом, а рострально переходит в большие полушария конечного мозга. На уровне промежуточного мозга водопровод расширяется и образует третий желудочек. Промежуточный мозг включает в себя несколько основных образований: зрительный бугор - таламус, подбугорье - гипоталамус, надбугорье - эпиталамус. С гипоталамусом структурно и функционально связана центральная железа внутренней секреции - гипофиз. Зрительный бугор, или таламус, формирует боковые стенки III желудочка и состоит из мощного скопления серого вещества. Зрительный бугор делят на собственно зрительный бугор и забугорную область или метаталамус. Основную массу серого бугра составляет таламус. В нем выделяют выпячивание — подушку, кзади от которой имеются два возвышения — наружное и внутреннее коленчатые тела (они в основном составляют забугорную область). Надбугорная область, или эпиталамус, состоит из шишковидной железы - эпифиза и задней спайки мозга. Эпифиз является железой внутренней секреции. Гормонпродуцирующими клетками шишковидной железы являются пинеалоциты, выделяющие днем серотонин, а ночью - мелатонин. Периодические колебания уровня мелатонина во многом определяют суточные биологические ритмы у высших животных и человека. Кроме того, отмечен антогонизм мелатонина с половыми гормонами, что во многом объясняет причины активизации репродуктивных функций организма в весенне-летний период. Забугорная область — метаталамус, состоящий из наружного и внутреннего коленчатых тел, имеет отношение к проведению зрительных (наружные коленчатые тела) и слуховых (внутренние коленчатые тела) афферентных импульсов. Зрительный бугор или таламус представляет собой парное скопление серого вещества промежуточного мозга. Нервные центры (ядра) зрительного бугра делят на три группы: специфические, ассоциативные и неспецифические. Специфические ядра зрительного бугра, получающие нервные импульсы от строго определенных афферентных зон и передающие эти импульсы в соответствующие ограниченные зоны коры (первичные проекционные зоны), называются проекционными или переключающими ядрами. К ним относятся вентролатеральные ядра. Переключающие ядра для зрительных и слуховых импульсов заложены соответственно в латеральных и медиальных коленчатых телах, прилежащих к задней поверхности зрительных бугров и составляющих основную массу забугорья. Спицефические ядра зрительного бугра принимают участие в анализе и переработке сигналов всех видов модальностей из внешней и внутренней среды перед тем, как они поступят в большие полушария. Неспецифические ядра являются представительством ретикулярной формации в зрительном бугре. Они рассматриваются как часть ретикулярной формации и объединяются под названием неспецифической диффузной таламической системы. Неспецифические ядра связаны с корой больших полушарий и структурами лимбико-ретикулярного комплекса. Эта система принимает участие в регуляции уровня возбудимости нейронов коры и играет определенную роль в сложном механизме формирования эмоций и соответствующих им выразительных непроизвольных движений, мимики, плача и смеха. Ассоциативные ядра, получая потоки нервных имульсов от специфических ядер, подвергают их обработке и направляют после этого к нейронам ассоциативной коры больших полушарий, где осуществляется формирование новых временных связей междк нервными центрами. Подбугорная область (гипоталамус) лежит книзу от зрительного бугра и представляет собой скопление высокодифференцированных ядер, которых насчитывают 32 пары. Все эти ядра разделяют на три группы: переднюю, среднюю, заднюю. Каждая группа ядер имеет свое функциональное значение. К среднему отделу ядер относятся серый бугор, воронка (инфундибулум) и нижний мозговой придаток — гипофиз. Подбугорная область является интегративным центром, посредством которого происходит сонастройка соматических, вегетативных и гуморальных мехнизмов регуляции. Определенные ядра гипоталамуса обладают нейросекреторными свойствами, т.е. выделяют вещества — гормоны, которые регулируют те или иные функции органов. Эти ядра тесно связаны с гипофизом — главной эндокринной железой организма. В нейронах гипоталамуса образуются гормоны, получившие название либеринов и стативно, которые, попадая в гипофиз, регулируют выделение тропных гормонов. Кроме того, в гипоталамусе синтезируются эффектрные гормоны окситоцин и вазопрессин, которые аккумулируясь в заднем отделе гипофиза, участвуют в регуляции водно-солевого обмена (вазопрессин или антидиуретический гормон) и активности миометрия матки (окситоцин). Гипоталамус контролирует деятельность всех эндокринных желез, более других — половых желез, щитовидной железы и надпочечников. Ядра подбугорной области принимают участие в регуляции всех видов обмена веществ и терморегуляции (т.е. в регуляции теплообмена организма). Структуры гипоталамуса является высшими интегративными центрами автономной (вегетативной) нервной системы: задние ядра гипоталамуса являются высшими интегративными центрами симпатического отдела вегетативной нервной системы, а передние ядра - парасимпатического отдела. В связи с чем, нервные центры гипоталамуса являются центрами, участвующими в регуляции вегетативного обеспечения деятельности внутренних органов и систем. Важная роль принадлежит гипоталамусу в регуляции сна. Поражение гипоталамуса может сопровождаться нарушениями сна и бодрствования. Гипоталамус является частью лимбической системы мозга и поэтому принимает участие в формировании эмоций и мотиваций лежащих в основе целенаправленного поведения высших животных и человека. Примитивные типы мотиваций (голод, жажда, сон, половое влечение) формируются при участии гипоталамуса. Он обеспечивает регуляцию вегетативных функций и осуществляет вегетативную окраску всех эмоций ЛЕКЦИЯ №12 МОРФО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ КОНЕЧНОГО План лекции 1. Анатомия конечного мозга 2. Физиология конечного мозга 3. Физиология лимбической системы Анатомия конечного мозга Конечный мозг развивается из переднего мозгового пузыря, состоит из хорошо развитых парных частей - правого и левого полушария и соединяющей их срединной части. Полушария разделены продольной щелью, в глубине которой лежит пластинка белого вещества, состоящая из волокон, соединяющих два полушария - мозолистое тело. Под мозолистым телом находится свод, представляющий собой два изогнутых волокнистых тяжа, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Спереди от столбов свода находится передняя спайка. Между передней частью мозолистого тела и сводом натянута тонкая вертикальная пластинка мозговой ткани - прозрачная перегородка. Полушарие образовано серым и белым веществом. В нем различают плащ, образованный лежащим по поверхности серым веществом - корой полушарий; обонятельный мозг и скопления серого вещества внутри полушарий - базальные ядра. Два последних отдела составляют наиболее старую в эволюционном развитии часть полушария. Полостями конечного мозга являются боковые желудочки. В каждом полушарии различают три поверхности: верхнебоковую (верхнелатеральную) выпуклую соответственно своду черепа, срединную (медиальную) - плоскую, обращенную к такой же поверхности другого полушария, и нижнюю - неправильной формы. Поверхность полушария имеет сложный рисунок, благодаря идущим в различных направлениях бороздам и валикам между ними - извилинам. Величина и форма борозд и извилин подвержены значительным индивидуальным колебаниям. Однако существует несколько постоянных борозд, которые ясно выражены у всех и раньше других появляются в процессе развития зародыша. Ими пользуются для разделения полушарий на большие участки, называемые долями. Каждое полушарие делят на пять долей: лобную, теменную, затылочную, височную и скрытую долю, или островок, расположенный в глубине боковой борозды. Границей между лобной и теменной долями является центральная борозда, между теменной и затылочной - теменно-затылочная. Височная доля отделена от остальных боковой бороздой (см.рис. 1)
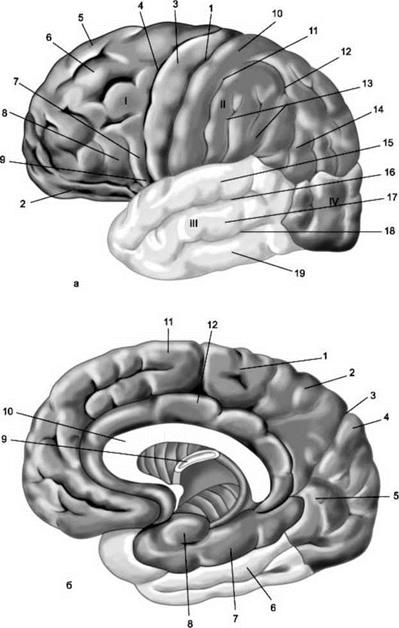
Рис. 1 Полушария большого мозга.
а - верхнелатеральная поверхность левого полушария: 1 - центральная борозда; 2 - глазничная часть нижней лобной извилины; I - лобная доля: 3 - прецентральная извилина; 4 - прецентральная борозда; 5 - верхняя лобная извилина; 6 - средняя лобная извилина; 7 - покрышечная часть нижней лобной извилины; 8 - нижняя лобная извилина; 9 - латеральная борозда; II - теменная доля: 10 - постцентральная извилина; 11 - постцентральная борозда; 12 - внутритеменная борозда; 13 - надкраевая извилина; 14 - угловая извилина; III - височная доля: 15 - верхняя височная извилина; 16 - верхняя височная борозда; 17 - средняя височная извилина; 18 - средняя височная борозда; 19 - нижняя височная извилина; IV - затылочная доля. б - медиальная поверхность правого полушария: 1 - парацентральная долька, 2 - предклинье; 3 - теменно-затылочная борозда; 4 - клин, 5 - язычная извилина; 6 - латеральная затылочно-височная извилина; 7 -
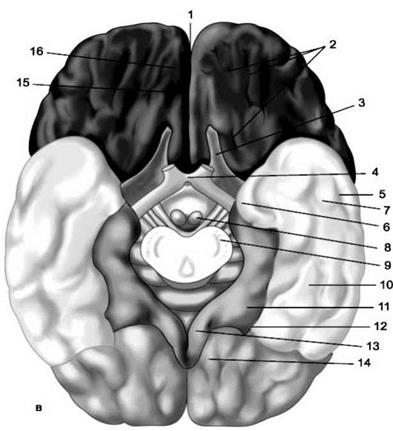
Рис. 2 Полушария головного мозга (нижняя поверхность)
в - нижняя поверхность больших полушарий: 1 - продольная межполушарная щель; 2 - глазничные борозды; 3 - обонятельный нерв; 4 - зрительный перекрест; 5 - средняя височная борозда; 6 - крючок; 7 - нижняя височная извилина; 8 - сосцевидное тело; 9 - основание ножки мозга; 10 - латеральная затылочно-височная извилина; 11 - парагиппокампальная извилина; 12 - коллатеральная борозда; 13 - поясная извилина; 14 - язычная извилина; 15 - обонятельная борозда; 16 - прямая извилина. На верхнелатеральной поверхности полушария (рис.1 а) в лобной доле различают предцентральную борозду, отделяющую предцентральную извилину, и две лобные борозды: верхнюю и нижнюю, делящие остальную часть лобной доли на верхнюю, среднюю и нижнюю лобные извилины. В теменной доле проходит постцентральная борозда, отделяющая постцентральную извилину, и внутритеменная, делящая остальную часть теменной доли на верхнюю и нижнюю теменные дольки. В нижней дольке выделяют надкраевую и угловую извилины. В височной доле две параллельно идущие борозды - верхняя и нижняя височные - делят ее на верхнюю, среднюю и нижнюю височные извилины. В области затылочной доли наблюдаются поперечные затылочные борозды и извилины. На медиальной поверхности (рис.1 б) хорошо видны мозолистое тело и поясная извилина. Над ней, окружая центральную борозду, лежит парацентральная долька. Между теменной и затылочной долями проходит теменнозатылочная борозда, а позади нее - шпорная борозда. Участок между ними называется клином, а лежащий впереди - преклиньем. В месте перехода на нижнюю (базальную) поверхность полушария лежит медиальная затылочновисочная, или язычная, извилина. На нижней поверхности (рис. 2), отделяя полушарие от ствола мозга, проходит глубокая борозда гиппокампа (борозда морского конька), кнаружи от которой находится парагиппокампальная извилина. Латеральнее она отделена коллатеральной бороздой от латеральной затылочно-височной извилины. Островок, расположенный в глубине латеральной (боковой) борозды, также покрыт бороздами и извилинами. Кора полушарий большого мозга представляет собой слой серого вещества толщиной до 4 мм. Она образована слоями нервных клеток и волокон расположенных в определенном порядке. Наиболее типично устроенные участки филогенетически более новой коры состоят из шести слоев клеток, старая и древняя кора имеет меньшее количество слоев и устроена проще. Разные участки коры имеют разное клеточное и волокнистое строение. В связи с этим существует учение о клеточном строении коры (цитоархитектоника) и волокнистом строении (миелоархитектоника) коры полушарии большого мозга.
|
|||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 378; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.27.244 (0.053 с.) |
 Структурно-функциональная организация продолговатого мозга;
Структурно-функциональная организация продолговатого мозга;