Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиология синаптической передачи
План лекции. 1. Понятие синапса. История развития представлений о синаптической передаче. 2. Классификация синапсов. 3. Современные представления о структуре химического синапса и механизмы передачи сигналов в нем. 4. Понятия о веществах синаптического действия. 5. Свойства химических синапсов. 6. Структура и свойства электрических синапсов. 1. Понятие синапса. История развития представлений о синаптической передаче. Синапсом называют образование, обеспечивающее передачу возбуждения от одной структуры к другой. Термин синапс (synapsis- греч. соединение, связь, застежка) ввел известный английский нейрофизиолог Чарльз Шерингтон (1897) для обозначения гипотетического образования или области, специализирую-щегося на обмене сигналами между клетками. Следует отметить, что в это время господствовала гипотеза о передачи информации с помощью биоэлектрических импульсов. Поэтому понятие, введеное Ч. Шерингтоном, обозначало место контакта между клетками, обеспечивающего передачу электрического импульса. Истоки представлений о возможной химической передаче возбуждения с нерва на мышцу уходят корнями к исследованиям великого французского физиолога К. Бернара (1850,1856). Исследуя механизм действия стрельного яда индейцев (кураре), К.Бернар обнаружил, что у животного, парализованного этим ядом, уменьшается как возбудимость мышц, так и проводимость иннервируемых эти мышцы нервов. На основании этих данных К. Бернар предположил, что кураре действует на нервные окончания, находящиеся в толще мышцы. Однако, исследования К. Бернара в то время не привели к формулировке представлений о химической природе передачи возбуждения в нервно-мышечном соединении. Впервые такую мысль сформулировал английский физиолог Т.Р. Элиот (1904). Эта гипотеза базировалась на сходстве действия адреналина на изолированное сердце и раздражение симпатических нервов. Однако, тем не менее не была воспринята его современниками. Прямое доказательство тому, что при раздражении блуждающих нервов выделяется химическое соединение, оказывающее аналогичное действие на изолированное сердце, было получено в работах австрийского фармаколога О. Леви (1921-1926). В утверждении принципа химической передачи в синапсах, большую роль сыграли работы российских ученых - А.Ф. Самойлова, А.В. Кибякова, А.Г. Гинецинского.
Исследуя температурную зависимость процесса перехода возбуждения с нерва на мышцу, А.Ф. Самойлов нашел, что она имеет высокий температурный коэффициент, что в большей степени подчеркивает химическую, а не физическую природу процесса распространения возбуждения. А.В. Кибяков (1933) предположил, что передача возбуждения с помощью химических веществ осуществляется не только в нервно-мышечных соединениях, но ив соединениях между нервными клетками. Разработав методику перфузии шейных ганглиев кошки, он показал, что если раздражать нервные волокна, подходящие к верхнему шейному ганглию, в последнем образуются химические вещества, способные вызвать возбуждения других ганглиев. Работами Гинецинского (1935) показано, что химические вещества в нервно-мышечных синапсах, взаимодействуя с эффекторными клетками, вызывают на небольшом участке мембран изменение мембранного потенциала, получившего название потенциала концевой пластинки. Наконец, решающую победу в химической природе передачи информации в синапсах принесли исследования английского физиолога Г. Дейла, который показал, что АХ не только имитирует активность парасимпатических отделов вегетативно-нервной системы, но и выделяется в нервных окончаниях нервно-мышечных синапсов. За серию работ в этой области в 1936 О. Леви и английскому физиологу Г. Дейлу была присуждена Нобелевская премия. Наконец, в связи с развитием электронной микроскопии в конце 50-х годов 20 столетия были открыты электрические синапсы, что подтвердило отчасти правоту Ч. Шерингтона. По поводу становления теории химической передачи информации в синапсе можно говорить очень много, однако, учитывая ограниченность отпущенного нам времени, перейдем к рассмотрению состояния вопроса на сегодняшний день. 2. Классификация синапсов. 1. По характеру расположения синапсы делят на центральные (в ЦНС) и периферические (в периферической нервной системе, синапсы на эффекторных органах). К центральным синапсам в зависимости от контактирующих
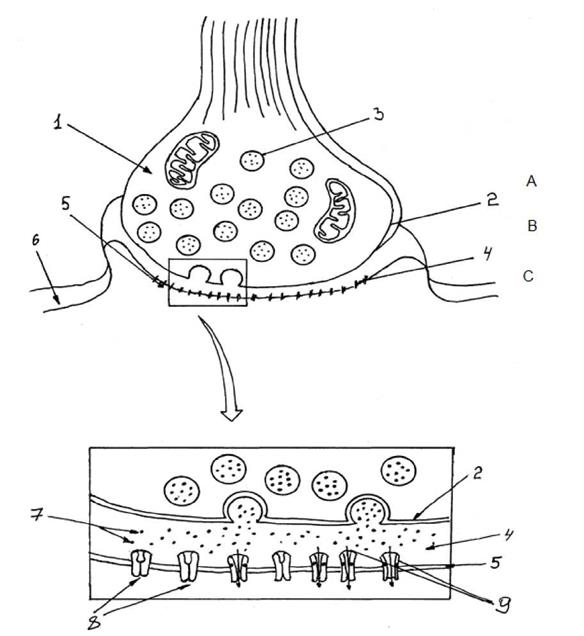
возбуждающих структур относят: - аксо-аксональные; - аксо-сомальные; - аксо-дендритные; - сома-дендритные; - денро-дендритные; - сома-сомальные. Периферические синапсы делят на: - нервно-мышечные синапсы скелетной поперечно-полосной мускулатуры; - нервно-мышечные синапсы гладкой мускулатуры; - нервно-мышечные синапсы миокарда; - синапсы вегетативных ганглиев; - нейросекреторные синапсы. В зависимости от механизма передачи синапсы делят на химические и электрические (нексусы миокарда, гладкой мускулатуры). Наконец, по характеру происходящих в постсинаптических образованиях процессов синапсы делят на возбуждающие и тормозящие. В зависимости от характера химического соединения участвующего в передаче сигнала через синапс их делят на холинэргические, адренэргические, серотонинэргические, гистаминэргические, пептидэргические и т.д. 3. Современные представления о структуре химического синапса и механизме передачи сигнала в нем. Данные электронной микроскопии, получившее развитие в последние десятилетия, позволили в химическом синапсе выделить 3 элемента (см.рис.1): A. пресинаптическую область (пресинаптическая терминаль); B. синаптическую щель; C. постсинаптическую область.
Рис. 1. Схема строения химического синапса. 1 - пресинаптическая область, 2 - пресинаптическая мембрана, 3 - везикулы содержащие нейротрансмиттер, A - пресинаптическая область (пресинаптическая терминаль), C - постсинаптическая область, 4 (B) - синаптическая щель; 5 - субсинаптическая мембрана, 6- постсинаптическая мембрана. 7- нейротрансмиттер в синаптической щели, 8 - эффекторные рецепторы, 9- хемочувствительный ионный канал.
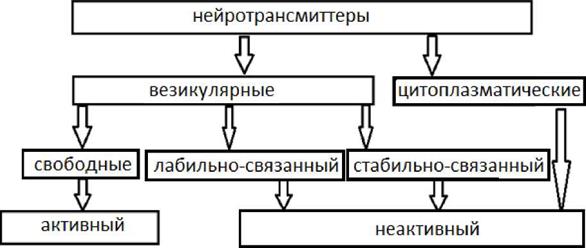
Пресинаптическая область представляет собой демиелинизированный терминальный участок отростка нервной клетки. По форме пресинаптическая терминаль напоминает собой колбу, прилежащую основанием к участку мембраны возбудимой клетки. Наиболее существенной чертой пресинаптической области является скопление пресинаптических пузырьков диаметром 50 мм (везикул), в которых содержится медиатор (химическое соединение, которое является материальным носителем сигнала к реципиентской клетке). Медиатор в пресинаптической области может находится в нескольких видах (см. рис.2).
Рис.2 Схема, отражающая формы локализации медиатора в пресинаптической области
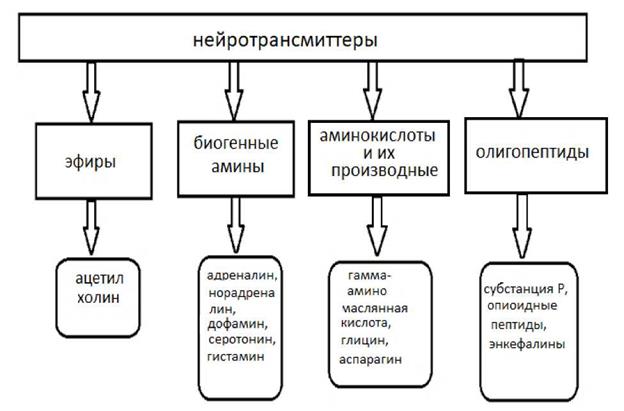
В пресинаптическом образовании содержатся белки, участвующие в обмене Са++ (кальмодулин и кальценейрин). Кальмодулин Са++ связывающие белки. Кальценейрин Кальмодулин - устойчивый белок М 17000, содержащий 4 Са++ связывающих центра. Кальцинейрин- антогонист кальмодулина. Ионы Mg++ могут ингибировать Са ++. Накопление Са++ может приводить к явлению посттетанической потенциации. В роли медиатора могут выступать различные химические соединения, которые можно разделить на ряд групп.
Рис. 3 Классификация медиаторов по химической структуре.
Кроме везикул с медиатора в пресинаптической области содержится большое количество митохондрий и лизосом, что свидетельствует о высокой активности обменных процессов в этой области. Кроме того, в этой области обнаружены предшественники медиаторов и продукты их метаболизма.
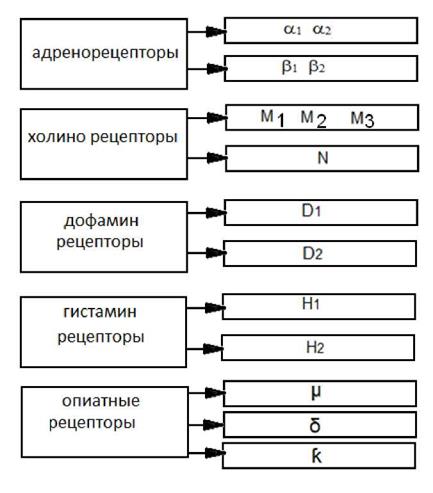
Синаптическая щель. В химических синапсах составляет от 20 до 50 мм. Здесь содержаться вода, электролиты, олигосахариды, ферменты, участвующие в расщеплении медиатора. Постсинаптическая область. Включает субсинаптическую мембрану (участок постсинаптической мембраны, имеющий специальный аппарат -рецепторы, характеризующиеся сродством к медиатору. В этой же области имеются химически-чувствительные ионные каналы. Собственно постсинаптическая мембрана - участок постсинаптической мембраны, которая содержит потенциалзависимые ионные каналы и на которой происходит генерация постсинаптических потенциалов. В зависимости от характера медиатора, с которым вступают во взаимодействие рецепторы субсинаптической мембраны, последние делятся на соответсвующие группы: 1) адренорецепторы (медиаторы норадреналин и адреналин); 2) холинорецепторы (медиатор ацетилхолин); 3) дофаминорецепторы (медиатор дофамин); 4) серотонинорецепторы (медиатор серотонин); 5) гистаминорецепторы (медиатор гистамин); 6) опиодидные рецепторы (медиаторы - эндогенные опиаты, энкефалины, эндорфины).
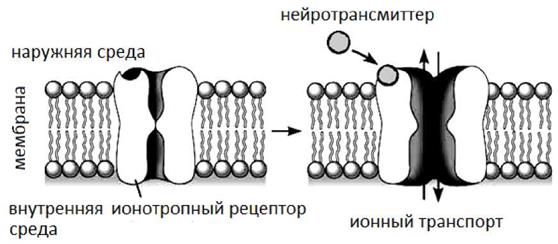
Следует отметить, что рецепторы субсинаптической мембраны классифицируются на ионотропные (лиганд-зависимые) рецепторы формирующие каналы и метаботропные рецепторы передающие сигнал от клетки к клетке с использованием G- белка и вторичных посредников. (см. Рис.5 и 6).
Рис. 5 Ионотропный рецептор, локализованный в мембране клетки.
физиологический ответ Рис.6 - Метаботропный рецептор, локализованный в мембране
До сравнительно недавнего времени считалось, что рецепторы, реагирующие на выделение медиатора, локализованы только в субсинаптической мембране. Однако, в последнее время они обнаружены и в пресинаптическом образовании. Рецепторные образования субсинаптической мембраны могут оказывать влияние на состояние ионных каналов субсинаптической мембраны. При этом это влияние может быть непосредственным (например, в некоторых холинэргических синапсах, так и через вторичных посредников). На сегодняшний день известен ряд соединений, которые могут выступать в роли вторичных посредников между рецепторами субсинап - тической мембраны и соответствующими биохимическими процессами клетки. 1) аденилат циклаза (АЦМФ); 2) гуанилат циклаза (ГЦМФ);
3) инозитол 3 фосфат; 4) фосфолипаза С; 5) Са++. Современные представления о механизмах передачи возбуждения в синапсе. 1) ПД распространяется по нервному волокну к пресинаптической области. 2) Изменение проницаемости мембраны пресинаптического образования к ионам Са++ и поступление Са++ в пресинаптическом образовании. 3) Движение везикул с активным медиатором пресинаптической области к пресинаптической мембране и выделение медиатора в синаптическую щель методом экзоцитоза. 4) Движение медиатора к субсинаптической мембране постсинаптической области и взаимодействие с соответствующими рецепторами мембраны. Далее возможны два варианта: 1- либо само взаимодействие медиатора с рецептором приводит к активации ионных каналов субсинаптической мембраны (это имеет место в некоторых холинэргических синапсах; 2 - либо вначале активируется вторичный посредник, а затем уже включается цепь биохимических процессов, приводящая к изменению проницаемости ионных каналов. Изменение проницаемости ионных каналов приводит к формированию местных токов и генерации на постсинаптической мембране постсинаптического потенциала. В зависимости от того по отношению к какому иону меняется проницаемость мембран, возможны два варианта постсинаптических потенциалов - возбуждающий, деполяризационный (ВПСП) и тормозной, гиперполяризационный (ТПСП). По своей сути указанные явления представляют собой локальные ответы. В основе ВПСП лежит повышение проницаемости мембраны по отношению к ионам Иа+, а ТПСП- по отношению к ионам К+ и Cl-. Характер активности тех или иных каналов определяется химической структурой медиатора, особенностью рецепторного образования, а также связанного с ним вторичного посредника. Например, в адренэргическом синапсе норадреналин может взаимодействовать либо с а адренореактивными образованиями, либо с р. Взаимодействие с а адренореактивными образованиями приводит к тому, что инозитол-3-фосфат активирует протеинкиназы, что через цепь промежуточных биохимических реакций приводит к повышению проницаемости ионных каналов по отношению к Na+, а это в свою очередь приводит к возникновению ВПСП. Если же норадреналин взаимодействует с р адренорецепорами в качестве вторичного посредника может выступать - аденилатциклаза - ЦАМФ, что приводит в конечном счете к повышению проницаемости ионных каналов по отношению к ионам К+, а это в свою очередь приводит к возникновению ТПСП. В ряде случаев имеются только тормозные медиаторы, взаимодействие которых с рецепторами приводит к повышению К+ проницаемости и возникновению ТПСП. К таким медиаторам относят гамма амино-маслянную кислоту и глицин. После взаимодействовия с рецептором, медиатор должен быть удален с мембраны. Это осуществляется ферментами, находящимися в синапсе. Ацетилхолин расщипляется ацетил-холинэстеразой. Для адреналина и норадреналина - такими ферментами является катехолокси-
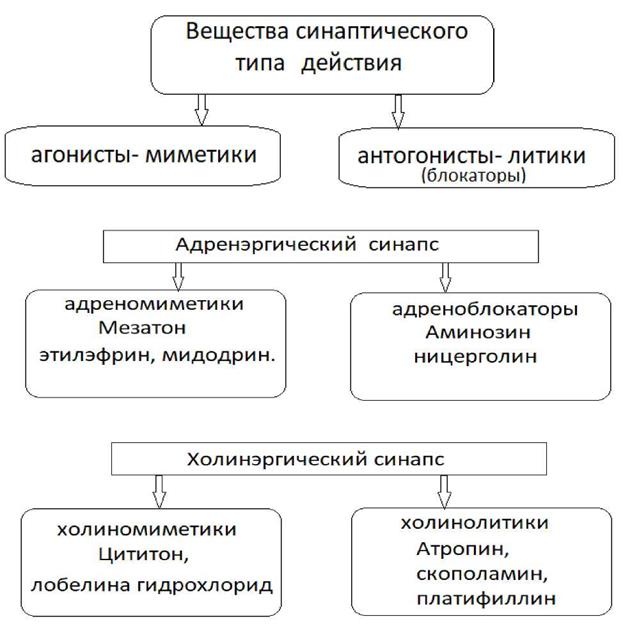
метилтрансфераза (КОМТ) и (МАО) моноаминоксидаза. Продукты расщепления медиаторов далее либо транспортируются в пресинаптическую область для последующего ресинтеза медиатора, либо удаляются из околосинаптической области. Регуляция активности синапсов. 1) Активность синапсов может регулироваться биологически активными веществами. Например, адренэргический синапс может тормозиться простогландином Е. 2) Активность синапсов может тормозиться медиаторами антогонистами Например, ацетилхолин тормозит адренэргический синапс. 4. Понятие о веществах синаптического действия. Ряд химических веществ могут воспроизводить или блокировать действие медиаторов. Вещества, воспроизводящие действие медиаторов получили название агонистов или меметиков. Вещества, блокирующие действие медиаторов получили название антагонистов медиаторов или литиков (см. рис. 7).
Рис.7. Вещества синаптического типа действия.
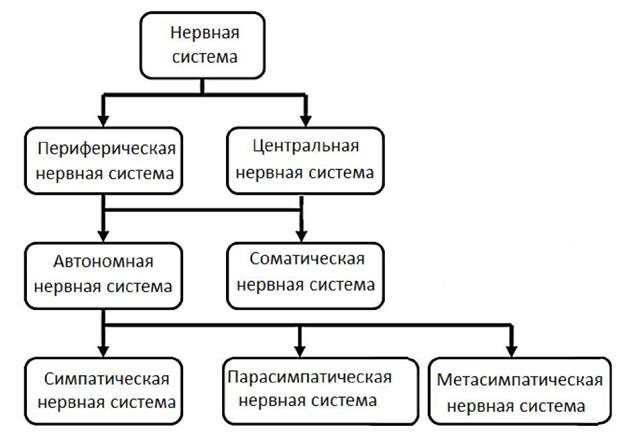
5. Свойства химических синапсов. 1. Одностороннее проведение возбуждения. 2. Способность к синаптической задержке. 3. Способность к суммации возбуждения на постсинаптической мембране. 4. Способность к трансформации возбуждения. 5. Способность к облегчению проведения возбуждения (посттетаническая потенциация). 6. Способность к актуализации синапса (проторение проведения возбуждения через синапс). 7. Повышенная утомляемость синапса. 8. Избирательная чувствительность к определенным химическим соединениям. 9. Низкая лабильность по сравнению с другими образованиями. 6. Структура и свойства электрических синапсов Электрические синапсы встречаются в организме животных и человека значительно реже химических. Электрические синапсы также как и химические имеют пресинаптическое образование, синаптическую щель и постсинаптическую мембрану. Отличительная особенность пресинаптического образования - отсутствие пузырьков с медиатором. Синаптическая щель значительно уже, чем у химических синапсов (у электрических синапсов - от 2 до 5нм, тогда как у химических синапсов - 2050 нм). Между пре- и постсинаптическими мембранами имеются каналы (щелевые контакты диаметром -1 нм. Данная структура имеет низкое электрическое сопротивление, что позволяет электрическому импульсу безпрепятсвенно переходить с одной клетки на другую. Следует добавить, что щелевые контакты не только электропроводны, но и проницаемы для низкомолекулярных метаболитов, обеспечивая единый химизм клеток, имеющих электрические синапсы. Поэтому электрические синапсы, как правило, встречаются у клеток, имеющих одно происхождение и единую специализацию. В отличии от химических, электрические синапсы проводят возбуждения в двух направлениях. У электрических синапсов отсутствует время задержки, слабая выраженность следовых процессов. Электрические синапсы характеризуются низкой надежностью в передаче информации. Свойства электрических синапсов. 1. Отсутствие центральной задержки. 2. Проведение возбуждения в обе стороны. 3. Относительно высокая лабильность. 4. Являются практически неутомляемыми образованиями. 5. Не чувствительны к химическим соединениям. 6. В электрических синапсах отсутствует явление посттетанической потенциации. 7. Более низкая надежность в передаче информации. Лекция №7 ОБЩАЯ МОРФОЛОГИЯ И ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ. План лекции 1. Общий план строения нервной системы. Общий план строения головного и спинного мозга. 2. Нейронная теория строения нервной системы. Строение и классификация нейронов. 3. Рефлекс как структурно-функциональная единица деятельности центральной нервной системы. История развития учения о рефлексе (Р.Декарт, И.Прохазка, И.М. Сеченов, И.П. Павлов, Н.А. Бернштейн, П.К.Анохин). 4. Принципы рефлекторной теории. 5. Классификация рефлексов. Процессы регуляции в организме животных и человека осуществляются нервными и гуморальными механизмами. Гуморальные механизмы регуляции связаны с влиянием на органы исполнители различных химических соединений через внутреннюю среду организма. Понятие внутренней среды организма ввел известный французский физиолог Клод Бернар. Под внутренней средой организма К.Бернар понимал совокупность биологических жидкостей организма - кровь, лимфу, тканевую жидкость. В качестве химических веществ участвующих в механизмах регуляции физиологических функций выступают биологически активные вещества, в т.ч. гормоны, электролиты и метаболиты (вещества образующиеся в ходе обмена веществ и энергии). Гуморальные механизмы регуляции характеризуются значительным латентным периодом, инертностью, высокой энергозатратностью, генерализованностью. Нервные механизмы характеризуются коротким латентным периодом, быстротой реализации, точной направленностью, низкой энергозатратностью. Сегодняшняя лекция будет посвящена общим представлениям о нервных механизмах регуляции физиологических функций. Нервную систему человека делят на центральную и перефирическую. Центральная нервная система включает спинной и головной мозг, а периферическая нервная система состоит из периферических нервов и узелков (ганглиев), в которых содержатся нервные клетки автономной (вегетативной) нервной системы. Головной мозг состоит из двух полушарий, промежуточного мозга, среднего мозга, продолговатого мозга, моста и мозжечка. Спинной мозг имеет сегментарное строение и представлен 5 (пятью) отделами: шейным, грудным, поясничным, крестцовым и копчиковым (см. рис. 1).
Рис. 1 Общий план строения центральной нервной системы.
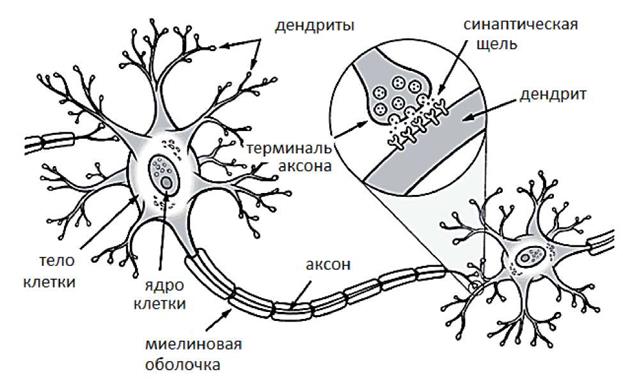
Более подробно о строении центральной нервной системы вы узнаете на лекциях и занятиях анатомии человека и на наших лекциях и занятиях, посвященных физиологии частных разделов центральной нервной системы. Кроме представленной выше классификации различных отделов нервной системы, существует и другая. Нервную систему, кроме того, делят на соматическую и автономную (вегетативную). Соматическая нервная система иннервирует скелетную поперечнополосатую мускулатуру и обеспечивает нервной регуляцией опорно-двигательный аппарат. Автономная нервная система иннервирует внутренние органы и обеспечивает их регуляцию в интересах организма. Автономная нервная система состоит из трех отделов: симпатического, парасимпатического и метасимпатического. Каждый из указанных отделов нервной системы имеет свою структурную организацию и конкретную функцию в организме. Нервная ткань, составляющая основу нервной системы, состоит из клеточных элементов и межклеточного вещества. Клеточные элементы нервной ткани. Основной клеточной единицей нервной ткани является нервная клетка - нейрон. Нервны клетки представляет из себя многоотросчатые образования размером от 4 до 130 мкм, среди которых выделяют отростки двух типов:
Рис. 2 Схема строения нейрона.
Ядро нейронов, как правило, округлой формы и находится в центре клетки. Наряду с другими органеллами в протоплазме нервных клеток имеются специализированные образования - нейрофибриллы, представляющие собой тонкие нити, состоящие из белка нейротензина, образующие в теле нейронов густую сеть. В протоплазме нейронов имеется большое количество рибосом. Необходимо отметить, что мемрана нейронов имеет неодинаковую возбудимость. В области отхождения от тела нейрона аксона существует участок мембраны, обладающий высокой возбудимостью. В этой части нейрона, как правило, и генерируются потенциалы действия, которые затем распространяются по аксону. Отростки нервных клеток, как правило, одеты оболочками и вместе с последними носят название нервных волокон. Различают две группы нервных волокон: миелиновые (мякотные) и безмиелиновые (безмякотные). Более подробно строение и классификацию нервных волокон вы будете изучать на кафедрах анатомии и гистологии. Классификация клеток нервной системы. 1. По количеству отростков все нейроны подразделяются на несколько групп: - группа нейронов с одним отростком (униполярные нейроны); - группа с двумя отростками (биполярные нейроны); - группа многоотросчатых нейронов (мультиполярные нейроны). В человеческом организме наиболее распространенными нейронами являются псевдоуниполярные нейроны (нейроны спинальных ганглиев) и мультиполярные нейроны (встречаются во всех отделах центральной нервной системы). 2. По выполняемым функциям нейроны делят на три группы: - чувствительные (афферентные); - вставочные (ассоциативные); - исполнительные (эффекторные). 3. По характеру вырабатываемого в нейронах медиатора нейроны делят на адренэргические, холинэргические, гистаминэргические, пептидэргические и т.д. 4. По характеру вызываемого ими эффекту нейроны делят на возбуждающие и тормозящие. 5. По форме нейроны делят на пирамидные, корзинчатые, звезчатые, овальные. В состав нервной ткани наряду с нейронами входят клетки нейроглии (глионы), которых в ЦНС в 5 - 10 раз больше, чем нейронов. Глионы различают по количеству имеющихся у них отростков: олигдендроциты (клетки с малым количеством отростков и астроциты (звезчатые клетки с большим количеством отростков. В периферической нервной системы глионы представлены так называемыми швановскими клетками. Олигодендроциты и швановские клетки формируют миелиновые оболочки вокруг осевых цилиндров нервных волокон. Наиболее распространенными глиальными клетками являются астроциты. Функции астроцитов: 1. Опорная, астроциты формируют «скелет» нервной системы; 2. Очищают межклеточное пространство нервной ткани от медиаторов и ионов (устраняют информационные помехи; 3. Выполняют защитную функцию (обладают фагоцитирующей активностью); 4. Способствуют трофике (питанию) наиболее активных клеток ЦНС; 5. Регулируют кровоток в нервной ткани; 6. Участвуют в формировании гематоэнцефалического барьера; 7. В последнее время показано, что глионы участвуют в механизмах формирования долговременной памяти. Рефлекс как структурно-функциональная единица деятельности Структурно-функциональной единицей деятельности центральной нервной системы является рефлекс. Первые представления о простом отражательном механизме окружающего мира, имеющемся у высших животных и у человека, впервые были сформулированы в XVII веке в работах известного французского богослова Рене Декарта (Р. Декарт служил монахом в монастыре). Взгляд Р. Декарта на человека характеризовался двойственностью. Простейшие реакции, например отдергивание руки от горячего предмета, по Р. Декарту можно представить в виде конкретного физиологического механизма, который обеспечивается нервами и мозгом и мышцами. Более сложные проявления нервной деятельности, например, высшие психические функции, согласно Р. Декарту связаны с богом, и в этой связи, они непознаваемы. Данные представления легли в основу одного из философских представлений о сущности человека, получившего название дуализма (двойственный взгляд на природу человека). Термин «рефлекс», обозначавший отражательный механизм нервной системы был введен несколько позже чешским ученым И. Прохазкой в 1784 году в «Трактате о функциях нервной системы». Представление о структуре рефлекторной в том виде, как мы ее представляем сегодня, по существу сформировалось в 19 столетии (работы Ч. Белла и Ф. Можанди). Более того в работах Ч. Белла мы находим уже представления не только о классической рефлекторной дуге, но и о рефлекторном кольце. Вклад российских ученых в развитие рефлекторной теории. Значительный вклад в развитие рефлекторной теории внесли представители Российской физиологической школы - И.М. Сеченов, И.П. Павлов, Н.А. Бернштейн, П.К. Анохин. И.М. Сеченов в 1863 году выпустил в свет книгу «Рефлексы головного мозга», в которой впервые попытался преодолеть дуализм Р. Декарта. Он высказал революционную для того времени мысль о том, что в основе всех проявлений деятельности центральной нервной системы, включая высшие психические явления, лежат рефлексы. Однако, как позже писал И.П. Павлов, это представление было сугубо «теоретическое, в виде физиологической схемы», не имевшее экспериментального физиологического обоснования. Павловский этап развития рефлекторной теории ознаменовался разработкой объективного метода изучения функций головного мозга, метода условных рефлексов. Этот метод позволил перевести ряд гипотез, выдвинутых И.М. Сеченовым, в плоскость экспериментально обоснованных теорий. Кроме того, И.П. Павлов сформулировал три основных принципа рефлекторной теории. Работы Н.Бернштейна и П.К.Анохина обогатили рефлекторную теорию представлением о механизме обратной связи. Таким образом, к середине 20 столетия сформировалось представление о рефлексе как механизме и структуре рефлекторной дуги (см.рис. 3).
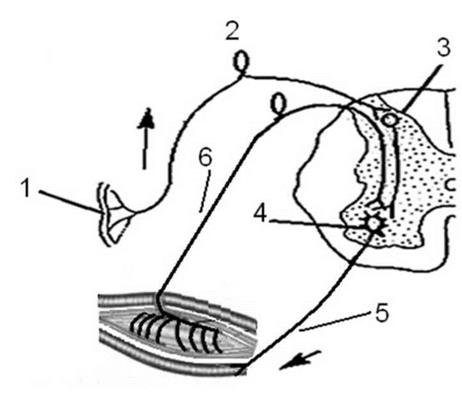
Рис. 3 Схема рефлекторной дуги спинального соматического рефлекса Обозначения: 1 - чувствительный рецептор; 2 - чувствительный (афферентный нейрон); 3 - вставочный ассоциативный нейрон; 4 - исполнительный (эффекторный) нейрон; 5 - исполнительный орган; 6 - обратная связь. Пунктирной линией обведен нервный центр. Рефлекс - это ответная реакция организма на раздражитель при участии центральной нервной системы в ответ на возбуждение рецепторов. Принципы рефлекторной теории: 1. Принцип детерминизма (принцип причинности). Этот принцип подчеркивает значимость раздражителя как фактора являющегося причиной возникновения рефлекса. Без действия раздражителя не может возникнуть рефлекторный ответ. 2. Принцип единства структуры и функции. Этот принцип подчеркивает единство структуры рефлекторной дуги и ее функции. При нарушении целостности любого звена рефлекторной дуги возникает нарушение рефлекса. 3. Принцип единства анализа и синтеза в рефлекторном механизме. Анализ - разделение целого на части. Синтез - формирование целого из отдельных частей. Простейший анализ в рефлексе реализуется на этапе взаимодействия рецептора с адекватным раздражителем. Говоря другим языком, рецептор из целостного окружающего мира «вычленяет» только те раздражители, взаимодействовать с которыми он приспособлен ходом эволюционного процесса. Рецептор преобразует энергию раздражителя в энергию универсального носителя информации в нервной системе (потенциал действия). Потенциал действия при помощи афферентного нейрона распространяется в нервный центр, где и осуществляется синтез, объединение различных сигналов. Под нервным центром понимают совокупность нейронов, расположенных в центральной нервной системе, которые обеспечивают реализацию того или иного рефлекса. 4. Принцип обратной связи. Этот принцип отражает роль обратной связи в реализации механизмов нервно-рефлекторной регуляции физиологических функций. Обратная связь - это механизм, обеспечивающий информацией о состоянии исполнительного органа нервные центры. Различают положительные и отрицательные обратные связи. Положительная обратная связь усиливает процесс, протекающий в нервном центре. Отрицательная обратная связь ослабляет процесс, протекающий в нервном центре. Классификация рефлексов 1. Классификация по критерию наследования. По этому критерию рефлексы делят на врожденные (видовые) и приобретенные (индивидуальные). Последнюю группу рефлексов И.П.Павлов назвал условными рефлексами. 2. Классификация по биологической значимости. По этому критерию рефлексы делят на пищевые, питьевые, оборонительные и т.д. 3. Классификация по тому, где в центральной нервной системе находится нервный центр рефлекса. По этому критерию рефлексы делят на рефлексы спинного мозга (спинальные), рефлексы продолговатого мозга (бульбарные), рефлексы среднего мозга (мезенцефальные) и т.д. 4. Классификация по характеристике отдела нервной системы, который формирует исполнительную часть рефлекса. По этому критерию рефлексы делят на соматические и вегетативные (автономные). 5. По времени протекания рефлексы делят на быстро реализующиеся (фазные) и длительно реализующиеся (тонические). 6. По характеристике исполнительного органа рефлексы делят на двигательные, секреторные, сердечные, сосудистые и т.д. 7. По отношению к раздражителю, вызвавшему рефлекс, рефлексы делят на положительные и отрицательные. Положительные рефлексы обеспечивают сближение с раздражителем. Отрицательные рефлексы обеспечивают, либо удаление от раздражителя, либо препятствуют сближению с ним. 8. Лекция № 8 НЕРВНЫЙ ЦЕНТР И ЕГО СВОЙСТВА. КООРДИНАЦИЯ План лекции. 1. Понятие «нервный центр». Две тенденции во взглядах на нервный центр. 2. Свойства нервных центров. 3. Координация в нервной системе, роль торможения в механизмах координации. 4. Виды торможения в центральной нервной системе. 5. Принципы координации. Исторически сложились два подхода к трактовке понятия нервный «центр»: узколокалистический и широкий. Узколокалистический подход к трактовке понятия «нервный центр» связан с работами известного французского физиолога М. Флуранса (1842) (M. Flourense, 1842), который показал, что нервный центр, обеспечивающий нервно-рефлекторную регуляцию дыхания у птиц находится в продолговатом мозге. Аналогичную точку зрения высказывал наш Российский ученый Н.А. Миславский, (1885). В соответствии с представлениями ученых, придерживающихся узколокалистического взгляда, «нервный центр» - это совокупность нейронов, локализованных в определенном участке центральной нервной системы и обеспечивающих реализацию конкретного рефлекса. Широкое толкование понятия «нервный центр» связано с работами российской физиологической школы (И.П. Павлов, 1923; П.К. Анохин, 1968 и некоторых других). В соответствии с представлениями ученых, придерживающихся этого направления, «нервный центр» - это совокупность нейронов, представленных на различных этажах центральной нервной системы, взаимосвязанных между собой для обеспечения регуляции какой- либо физиологической функции. В качестве примера рассмотрим «дыхательный нервный центр». Исполнительными органами в системе внешнего дыхания являются дыхательные мышцы, обеспечивающие изменение объема грудной клетки. К таким мышца относят диафрагму и наружные косые межреберные мышцы. Мотонейроны, иннервирующие данные группы мышц находятся в спинном мозге. Следовательно, обсуждая понятие «дыхательный нервный центр» мы должны упоминать Данные группы нейронов. Вместе с тем, мотонейроны спинного мозга находятся под контролем нейронов «собственно дыхательного центра», находящегося в продолговатом мозгу. Мотонейроны продолговатого мозга, водящие в состав «дыхательного нервного центра», в свою очередь контролируются нейронами Варолиева моста («пневмотаксический дыхательный центр»), гипоталамических центров, центров коры больших полушарий. Перечисленные выше нервные центры различных уровней ЦНС находятся во взаимодействии, иерархически организованы, оптимально обеспечивая регуляцию дыхания. Первый подход к трактовке понятия «нервный центр» в большей степени используется в неврологии, для топической диагностики повреждения тех или иных нервных центров. Второй подход используют специалисты в области физиологии для объяснения механизмов регуляции физиологических функций. Однако, не зависимо от варианта определения нервный центр - совокупность взаимосвязанных между собой нейронов, имеющих множество синапсов, различные физиологические свойства, коллатерали и «нейронные ловушки». Данные обстоятельства и привели к формированию у нервных центров множество разнообразных свойств. Свойства нервных центров. 1. Одностороннее проведение возбуждения. Данное свойство обусловлено наличием большого количества синаптических контактов, а синапсы проводят возбуждение только в одном направлении. 2. Наличие в синапсах феномена центральной задержки проведения возбуждения. Это свойство также обусловлено наличием в нервном центре большого количества синапсов.
|
|||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 382; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.127.141 (0.104 с.) |