Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности гладкомышечной ткани
Гладкомышечные клетки распределены в стенках внутренних органов - сосудах, органах желудочно-кишечного тракта, мочевом пузыре, мочевыводящих путях, матке. Гладкомышечные клетки имеют веретенообразную форму длиной от 50 до 400 микрон и толщиной от 2 до 10 микрон и содержат одно ядро. Сократительные элементы также, как и в скелетной мускулатуре представлены миофиламентами, содержащими актин, миозин и регу-ляторные белки. Однако, в отличие от скелетной мускулатуры в гладкомышечной ткани миофиламенты расположены беспорядочно, что и не приводит к возникновению поперечной исчерченности. В гладкомышечных клетках слаборазвит саркоплазматический ретикулум, поэтому ионы Са++ депонируются в цитоплазме клеток. Особенностью миозина гладкомышечной ткани является его низкая АТФазаная активность, что приводит к медленному скольжению нитей актина и миозина по отношению друг друга. В этой связи гладкомышечная ткань характеризуется медленной, тонической формой мышечного сокращения.
Рис. 2 Схема Т-системы скелетного мышечного волокна Кроме того, это приводит к низкой энергозатратности сокращения, поэтому в гладкомышечной ткани утомление развивается медленнее, чем в поперечнополосатой ткани скелетных мышц. Между отдельными клетками гладкомышечной ткани имеются специальные образования - нексусы (разновидности электрических синапсов), позволяющие возбуждению в виде биоэлектрического процесса достаточно быстро распространяться по ткани. Следует добавить, что гладкомышечные клетки способны переходить из состояния относительного покоя в состояние возбуждения без внешних воздействий, т.е. обладают свойством автоматии. Сравнительная характеристика скелетных и гладких мышц представлена в таблице 1. Таблица 1
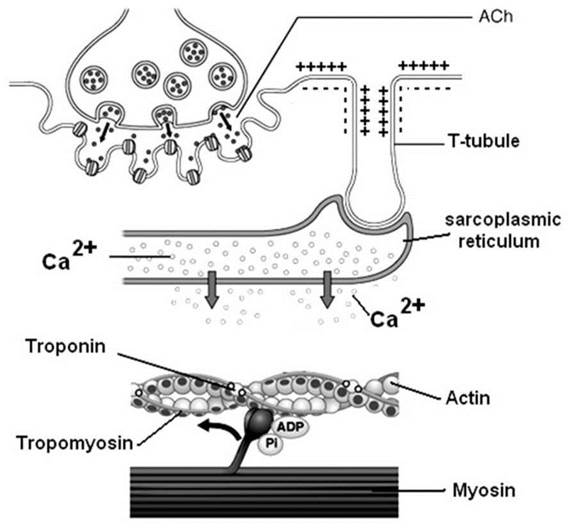
2. Двигательная единица. Классификация. Поперечнополосатая скелетная мышечная ткань иннервируется соматической нервной системой. Нейроны соматической нервной системы, иннервирующие скелетную мускулатуру, называются мотонейро-нами. Мотонейроны, иннервирующие белые мышечные волокна называются альфа! мотонейронами (а1-мотнейроны), мотонейроны, иннервирующие красные мышечные волокна называются альфа2 мотонейронами (а2-мотнейроны). Один мотонейрон, как правило, иннервирует достаточно много мышечных волокон (от 12 до 2500). Вместе с иннервирующим их мотонейроном мышечные волокна составляют двигательную или нейромоторную единицу. Двигательные единицы, участвующие в обеспечении тонких движений содержат от 12 до 15 мышечных волокон. К таким нейромоторным единицам относятся, например, двигательные единицы мышц кистей рук. Двигательные единицы, участвующие в обеспечении грубых движений содержат от 1500 до 2500 мышечных волокон. К таким двигательным единицам относят, например, нейромоторные единицы мышц нижних конечностей, глубоких мышц спины. В соответствии с составом двигательных единиц их делят на 3 группы:
I - медленные, неутомляемые двигательные единицы; IIA - быстрые, устойчивые к утомлению двигательные единицы; 11Б - быстрые, легко утомляемые двигательные единицы. I - медленные, неутомляемые двигательные единицы. Состоят из мелких а2-мотнейронов и красных, тонических мышечных волокон. Данные двигательные единицы характеризуются высокой возбудимостью, т.е. данные нейромоторные единицы переходят в состояние возбуждения при относительно низкой частоте разрядов потенциалов действия нервных клеток. Красные мышечные волокна, входящие в состав данных двигательных единиц, характеризуются хорошо развитой капилярной сетью, что позволяет им поддерживать длительное возбуждение. IIA - быстрые, устойчивые к утомлению двигательные единицы. Состоят из а1-мотнейронов и белых мышечных волокон. Также как и в предыдущей группе характеризуются хорошо развитой капилярной сетью. Мышечные волокна для воспроизводства энергии используют как аэробные, так и анаэробные процессы. По уровню возбудимости относятся к группе средневозбудимых двигательных единиц. 11Б - быстрые, легко утомляемые двигательные единицы. Состоят из а1-мотнейронов и белых мышечных волокон. Из представленных трех групп двигательных единиц характеризуется самой низкой возбудимостью и вместе с тем, максимальной амплитудой мышечного сокращения. Биохимической особенностью данной группы двигательных единиц является способность их мышечных волокон извлекать энергию для своих нужд за счет анаэробных процессов, т.е. без участия кислорода. Из представленной выше классификации следует, что нейромоторные или двигательные единицы обладают неодинаковой возбудимостью. Это обстоятельство приводит к тому, что при увеличении силы раздражителя, наносимого на мышцу количество, вовлеченных в процесс возбуждения двигательных единиц увеличивается, что закономерно приводит к увеличению амплитуды мышечного сокращения. Данная закономерность называется «Законом силы». «Закон силы» формулируется следующим образом: «Амплитуда сокращения скелетных мышц зависит от силы раздражителя: в определенном диапазоне с увеличением силы раздражителя возрастает амплитуда мышечного сокращения». 3. Современные представления о механизме мышечного сокращения. Причиной возбуждения мышечных волокон скелетной мышцы является потенциал действия, распространяющийся в рамках нейро-моторной единицы к нервно-мышечным синапсам. Следствием этого является возникновение возбуждающих постсинаптических потенциалов на постсинаптических мембранах нервно-мышечных синапсов. При определенных условиях указанные процессы трансформируются в потенциалы действия мышечных волокон, которые распространяются без затухания по биологической мембране, попадая, в том числе в область Т- системы. Следствием этого является изменение проницаемости потенциал зависимых ионных кальциевых каналов цистерн саркоплазматического ретикулума Т-системы и выход Са++ в саркоплазму. Ионы кальция выступают в роли фактора электромеханического сопряжения биоэлектрических процессов протекающих в области биологической мембраны мышечного волокна и механическим процессом скольжения нитей актина и миозина по отношению друг к другу. Точкой приложения ионов кальция являются регуляторные белки тропонин и тропомиозин, которые под воздействием ионов кальция изменяют свою конфигурация и деблокируют активные участки актино-миозиновых мостиков, взаимодействие между которыми и приводит к укорочению мышечного волокна. Следует добавить, что при открытие активных участков актина и миозина происходит расщепление молекул АТФ, поскольку активные участки миозина обладают АТФ-азной активностью. В результате расщепления АТФ выделяется энергия, которая расходуется в меньшей степени на процесс взаимодействия актиновых и миозиновых нитей, т.е. на укорочение мышечного волокна, но в большей степени на процесс на работу кальциевого насоса, удаляющего ионы кальция из зоны взаимодействия с регуляторными белками в саркоплазматический ретикулум с последущим депонированием в цистернах Т-системы. В связи с этим наступает процесс расслабления мышечных волокон. Данное обстоятельство подчеркивает, что процесс расслабления мышечной ткани более энергозатратный чем, процесс укорочения мышечных волокон.
4. Виды и режимы мышечного сокращения. Выделяют три режимы мышечного сокращения: 1. Изотонический; 2. Изометрический; 3. Смешанный (ауксометрический). 1. Изотонический режим мышечного сокращения характеризуется преимущественным изменением длины мышечного волокна, без существенного изменения напряжения. Указанный режим мышечного сокращения наблюдается, например, при поднятии легких и средних по массе грузов. 2. Изометрический режим мышечного сокращения характеризуется преимущественным изменением мышечного напряжения, без существенного изменения длины. Примером может служить изменения состояния мышц при попытке человека сдвинуть с места предмет большой массы (например, при попытке сдвинуть с места стену в комнате). 3. Смешанный (ауксометрический) тип мышечного сокращения, наиболее реальный, наиболее часто встречающийся вариант. Содержит в себе компоненты первого и второго вариантов в разных соотношениях в зависимости от реальных условий окружающей среды. Виды мышечного сокращения Выделяют три виды мышечного сокращения: 1. Одиночное мышечное сокращение; 2. Тетаническое мышечное сокращение (тетанус); 3. Тоническое мышечное сокращение. Кроме того, тетаническое мышечное сокращение делят на зубчатый и гладкий тетанус.
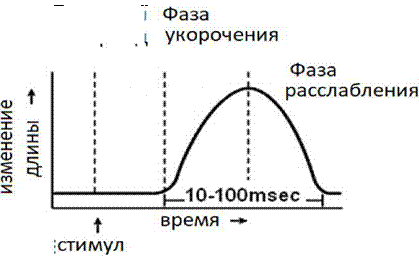
1. Одиночное мышечное сокращение возникает в условиях действия на мышцу пороговых или надпороговых электрических стимулов, межимпульсный интервал которых равен или больше длительности одиночного мышечного сокращения. В одиночном мышечном сокращении выделяют три временных отрезка: латентный период, фазу укорочения и фазу расслабления (см. рис. 3).
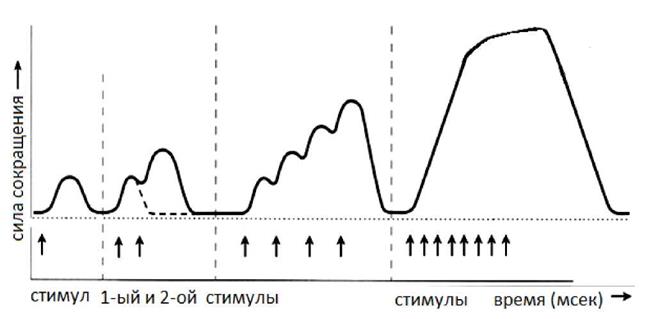
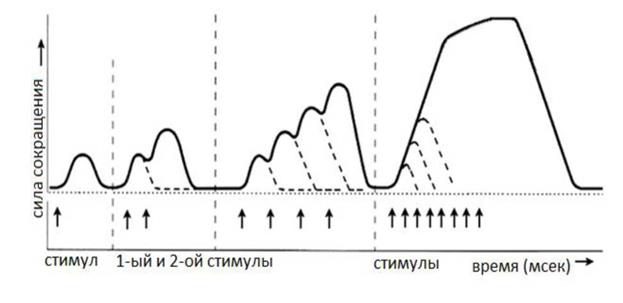
2. Тетаническое мышечное сокращение (тетанус) возникает в условиях действия на скелетную мышцу порогового или надпорогового электрического раздражителя, межимпульсный интервал которого меньше длительности одиночного мышечного сокращения. В зависимости от длительности межстимульных интервалов электрического раздражителя при его воздействии может возникнуть либо зубчатый, либо гладкий тетанус. Если межимпульсный интервал электрического раздражителя меньше длительности одиночного мышечного сокращения, но больше или равен сумме латентного периода и фазы укорочения, возникает зубчатый тетанус. Указанное условие выполняется при повышении частоты импульсного электрического раздражителя в определенном диапазоне. Если же длительность межимпульсного интервала электрического раздражителя меньше суммы латентного периода и фазы укорочения возникает гладкий тетанус. При этом амплитуда гладкого тетануса больше амплитуды и одиночного мышечного сокращения и зубчатого тетанического сокращения. При дальнейшем уменьшении межимпульсного интервала электрического раздражителя, а следовательно при увеличении частоты, амплитуда тетанических сокращений возрастает (см. рис. 4).
Рис. 4 Зависимость формы и амплитуды тетанических сокращений от частоты раздражителя. Однако, указанная закономерность не носит абсолютного характера: при определенном значении частоты вместо ожидаемого повышения амплитуды гладкого тетатнуса отмечается феномен ее снижения (см. рис. 5). Указанный феномен был впервые обнаружен Российским ученым Н.Е. Введенским и был назван пессимумом. В основе пессимальных явлений по мнению Н.Е. Введенского лежит механизм торможения.
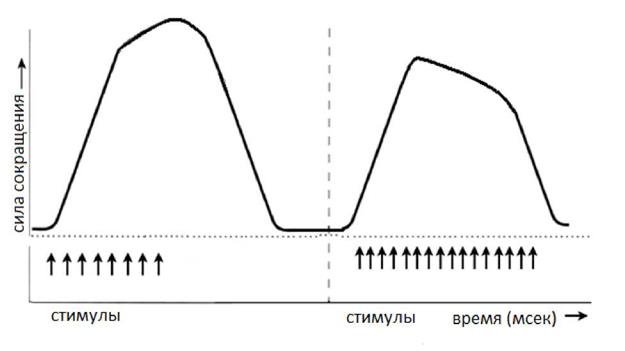
Рис. 5. Зависимость амплитуды гладкого тетануса от частоты раздражителя. Обозначения те же, что и на рисунке 5.
Механизмы тетанического сокращения Одной из наиболее распространенных теорий тетанического сокращения является теория предложенная в 19 веке известным немецким физиологом Гермоном Гельмгольцем. Эта теория получила название «Теория суперпозиции» («Теории суммации»). Согласно представлениям Г.Гельмгольца тетанус возникает в результате суммации механических проявлений изменения длины скелетных мышц. Если суммация последующего изменения длины мышцы с предыдущим происходит в условиях, когда мышечное волокно находится в фазе расслабления, возникает зубчатый тетанус. Если же суммация происходит в условиях попадания каждого последующего стимула на мышечное волокно, находящееся в фазе укорочения, возникает гладкий тетанус (см. рис. 6).
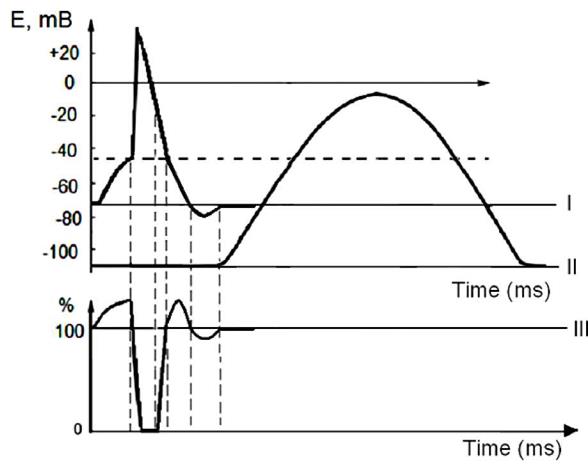
Рис. 6. Суммация мышечных сокращений при возникновении тетануса Физиологическая основа возможности суммации мышечных сокращений при действии относительно высокочастотных электрических раздражителей становится ясной при рассмотрении соотношений между изменением длины мышечного волокна в условиях одиночного мышечного сокращения, изменением мембранного потенциала и ее возбудимости (см. рис. 7).
Рис. 7 Соотношение между изменением длины мышечного волокна в условиях одиночного мышечного сокращения, изменением мембранного потенциала и ее возбудимости. Как следует из представленных соотношений, колебание мембранного потенциала при возбуждении мышечного волокна завершается в течение латентного периода одиночного мышечного сокращения. К этому же моменту мышечное волокно полностью восстанавливает свою возбудимость. В связи с эти оно способно полноценно возбуждаться, проявлением чего является полноценное дополнительное изменение длины мышечного волокна, которое может суммироваться с предыдущим. 5. Утомление в мышечной ткани Утомление - это временное состояние возбудимых образований, которое возникает в результате длительной или интенсивной работы и связано с истощением энергетических и пластических ресурсов, которые необходимы для возбуждения и специфической деятельности. Для устранения данного состояния необходимо восстановление ресурсов возбудимых образований, которое требует времени. В связи с истощением энергетических ресурсов в возбудимых тканях нарушается работа энергозатратных транспортных механизмов, что приводит к накоплению в тканях продуктов метаболизма, нарушению ионной асимметрии, характеризующей работоспособную клетку. В мышечной ткани утомление характеризуется снижением количества АТФ, увеличением количества АДФ, остатков фосфорной кислоты, молочной кислоты, ионов СА++. Утомление в скелетных мышцах проявляется в увеличении латентного периода, увеличением фазы расслабления, а в ряде случаев в возникновении остаточного укорочения (контрактуры), которое связано с накоплением в саркоплазме Са++ и затрудненим разобщения активных участков актина и миозина. Лекция №6
|
||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 256; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.145.114 (0.033 с.) |