Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Отличия соматического и вегетативного рефлексов
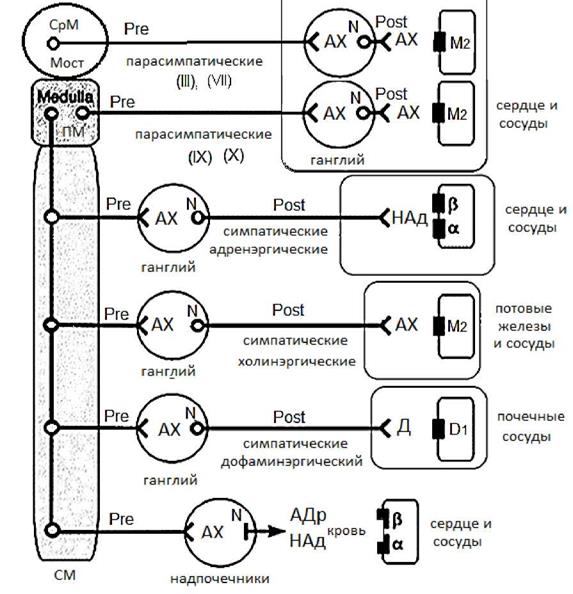
1. Расположение вставочного и исполнительного (эффекторного) нейронов. В соматическом рефлексе ассоциативная и исполнительная клетки расположены в сером веществе центральной нервной системы (в нашем случае в сером веществе спинного мозга). В вегетативном рефлексе первая клетка (ассоциативная) расположена в сером веществе центральной нервной системы, вторая - в ганглии (узелке) за пределами центральной нервной системы. Следствием этого является различные последствия перерезки передних корешков для соматических и вегетативных нервов. Соматические нервы дегенерируют, перерождаются, тогда как вегетативные нервы не перерождаются, а продолжают выполнять свои функции. 2. Места выхода нервных волокон за пределы ЦНС. Соматические волокна покидают ствол мозга, спинной мозг преимущественно сегментарно и перекрывают иннервируемые органы в пределах трех смежных сегментов. Волокна автономной нервной системы покидают центральную нервную систему в пределах трех относительно изолированных областях: ствол мозга (средний и продолговатый мозг), грудной, поясничный и крестцовый отделы спинного мозга. 3. Распределение волокон соматического и вегетативного нервов на периферии. Волокна соматических нервов на периферии распределяются преимущественно сегментарно, тогда как волокна вегетативных нервов распределяются преимущественно деффузно. 4. Морфологическая характеристика соматических и вегетативных волокон. Нервные волокна соматической нервной системы в основном покрыты миелином, в связи с чем, скорость распро-странения возбуждения в них достигает 100 - 120 м/сек. Нервные волокна вегетативной нервной системы либо не имеют миелина, либо содержат его в значительно меньшем количестве, чем нервные волокна соматической нервной системы. В этой связи скорость распространения возбуждения значительно меньше: 0,5 м/сек - 2 м/сек. Существенные морфологические и нейрохимические отличия имеют симпатический и парасимпатический отделы автономной нервной системы. Данные отличия выявляются при сравнении эффекторных частей данных отделов автономной нервной системы (см. рис. 2 и таблицу 1). Таблица 1 СРАНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СИМПАТИЧЕСКОГО И ПАРАСИМПАТИЧЕСКОГО ОТДЕЛОВ АВТОНОМНОЙ НЕРВНОЙ СИСТЕМЫ
Рис. 2 Схема организации эффекторных частей вегетативной нервной системы организма. Обозначения: СМ спинной мозг; ПМ - продолговатый мозг; АДр- адреналин; N- никотин чувствительные рецепторы; М - мускарин чувствительные рецепторы; Ах - ацетилхолин; НАд - норадреналин; III, VII, IX и X пары черепно-мозговых нервов (III - nervus oculomotorius; VII - nervus facialis; IX - nervus glossofaringius; X - nervus vagus)
Синаптическая организация автономной нервной системы Ганглионарные синапсы Синапсы ганглиев как симпатического, так и парасимпатического отделов в качестве медиатора используют ацетилхолин, т.е. являются холинэргическими. Эффекторные рецепторы постсинаптических мембран чувствительны не только к ацетилхолину, но и алколоиду, выделенному из листьев табака никотину. (Алколоиды - азотсодержащие органические соединения природного растительного происхождения, обладающие свойством оснований.) Поэтому они называются никотин чувствительными или Н - репторами. Особенностью данных рецепторов является то, что они блокируются ганглиоблокаторами (бензогексонием, пириленом). Физиологические особенности ганглионарных синапсов:
1) Значительная синаптическая задержка проведения возбуждения, в 5 раз превышающая по длительности аналогичный показатель центральных синапсов; 2) Большая длительность постсинаптических потенциалов; 3) Выраженная и длительная гиперполяризация. Эффекторные синапсы Эффекторные синапсы парасимпатического отдела автономной Эффекторные синапсы парасимпатического отдела автономной нервной системы являются холинэргическими, т.е. в роли медиатора в данных синапсах выступает ацетилхолин. Эффекторные рецепторы постсинаптических мембран данных синапсов чувствительны не только к ацетилхолину, но и к растительному алколоиду, выделенному из красных мухоморов - мускарину. Поэтому эффекторные рецепторы в данных синапсах называют мускарин чувствительными или М - рецепторами. М - рецепторы блокируются препаратами типа атропина. В зависимости от вторичного посредника, обеспечивающего связь активированного рецептора с цитоплазматическими процессами и характера вызываемого в клетке процесса, М-рецепторы делят на М1 и М2. Взаимодействие ацетилхолина с М1 вызывает в клетке возбуждение, тогда как активация М2 - рецепторов вызывает торможение. Эффекторные синапсы симпатического отдела автономной нервной В роли медиаторов эффекторных синапсов симпатического отдела в основном выступают норадреналин и адреналин. Поэтому эти синапсы называются адренэргическими. Эффекторные рецепторы постсинап-тической мембраны делят на а и 0. При взаимодействии медиатора с а адренорецепторами в эффекторной клетке возникает возбуждение, а при взаимодействии медиатора с 0 адренорецептором - торможение. Функциональный антагонизм симпатического и парасимпатического Основная масса внутренних органов имеет двойственную иннервацию: симпатическую и парасимпатическую. В соответствии с истори-чески сложившимся взглядом симпатический и парасимпатические отделы автономной нервной системы находятся в реципрокных отношениях. Согласно этому взгляду повышение активности одного отдела автономной нервной системы сопровождается торможением другого. При этом, как считал американский физиолог Уолтер Кэннон, активация симпатического отдела автономной нервной системы приводит к повышению деятельности физиологических систем организма и повышению уровня обмена веществ и энергии. Указанные сдвиги возникают в условиях действия на организм различных раздражителей, в т.ч. экстремальных и обеспечивают его приспособление к изменяющимся условиям. В комплексе данные измения получили название эрготропных процессов. В противоположность процессам эрготропного характера, повышение активности парасимпатического отдела автономной нервной системы связано с понижением энергетического обмена организма и с переориентацией его на анаболические, запасательные процессы. Другими словами основной функцией парасимпатического отдела автономной нервной системы является восстановление ресурсов организма, растраченных в ходе активного приспособления к изменяющимся условиям окружающей среды. Указанные процессы получили название трофотропных. С некоторой долей условности активную часть суток (день) можно назвать временем доминирования симпатического отдела автономной нервной системы, а ночное время, когда человек спит - временем «царствования» парасимпатического отдела автономной нервной системы. Рассмотрим примеры функционального состояния человека либо при доминировании симпатического отдела автономной нервной системы, либо парасимпатического ее отдела.
Доминирование симпатического отдела автономной нервной системы Наблюдается при воздействии на организм сильных и сверхсильных раздражителей. В условиях повышения тонуса симпатического отдела автономной нервной системы отмечается увеличение диаметра зрачков, увеличение частоты и силы сердечных сокращений, увеличение частоты и глубины дыхательных движений, расширение бронхов, перераспределение тонуса сосудов (коронарные сосуды, сосуды скелетных мышц расширяются, периферические сосуды кожи, сосуды желудочно-кишечного тракта суживаются). Повышается уровень артериального давления, снижается секреторная и двигательная активность желудочно-кишечного тракта, повышается свертываемость крови, увеличивается количество глюкозы в крови. Все указанные изменения способствуют приспособлению организма при воздействии на него различных раздражителей, в т.ч. экстремальных. Доминирование парасимпатического отдела автономной нервной Наблюдается в условиях относительного физиологического покоя, после прекращения действия на организм раздражителей, в отдельные фазы сна. В условиях повышения тонуса парасимпатического отдела автономной нервной системы отмечается уменьшение диаметра зрачков, уменьшение частоты и силы сердечных сокращений, сужение просвета бронхов, снижение частоты и глубины дыхательных движений, снижение уровня артериального давления. Кроме того, отмечается активизация желудочнокишечного тракта, снижение количества глюкозы в крови, понижение свертываемости крови. Все указанные изменения направлены на восстановление растраченных ранее резервов. Структурно-функциональная организация метасимпатического отдела Многими исследователями показано, что извлеченные из организма мышечные полые органы, в соответствующей питательной среде и определенном температурном режиме продолжают выполнять присущие им функции без видимых изменений. Например, деятельность изолированного сердца лягушки, перистальтика изолированной кишки. Эта функциональная автономия во многом объясняется наличием в стенках внутренних органов ганглиозной системы, которая наряду с другими механизмами участвует в регуляции функциональной активности данных органов. Указанная система ганглиев, находящаяся в стенках внутренних полых органов, рассматривается как относительно самостоятельный отдел автономной нервной системы. В России, благодаря работам А.Д. Ноздрачева, этот отдел называют метасимпатическим отделом автономной нервной системы.
Характеристика нейронов метасимпатического отдела автономной Исследованиями российского физиолога А. Догеля показано, что в интрамуральных ганглиях различают три типа нейронов: 1) I тип нейронов - исполнительные, эффекторные нейроны. Нейроны с большим количеством дендритов и аксоном, не имеющим миелиновой оболочки. Аксоны клеток I типа заканчиваются на гладкомышечных клетках; 2) II тип нейронов - чувствительные, афферентные нейроны. Основная особенность - не имеют на дендритах и соме синаптических контактов. Аксоны клеток II типа в пределах ганлиев контактируют с клетками III типа; 3) III тип нейронов - ассоциативные, вставочные нейроны. Обеспечивают взаимодействие нейронов II и I типа. Синаптическая организация метасмпатического отдела автономной В целом нейрохимическая организация синапсов метасимпатической нервной системы аналогична синаптической организации симпатического и парасимпатического отделов: встречаются холинэргические и адренэргические синапсы. Однако, в последние годы обнаружены синапсы, в которых функцию медиатора выполняет АТФ. Такие синапсы получили название пуринэргических и по своей функции являются тормозными. Таким образом, наличие трех функциональных классов нейронов в интрамуральных ганглиях позволяет им осуществлять нейрвно- проводниковые механизмы регуляции, напоминающие рефлексы. Функции метасимпатического отдела автономной нервной системы 1) Метасимпатический отдел автономной нервной системы выступает как самостоятельное образование, обеспечивающее нервную регуляцию на местном уровне; 2) Метасимпатический отдел автономной нервной системы может выступать как исполнительный регуляторный механизм, обеспечивающий реализацию влияние вышележащих отделов центральной нервной системы, в т. ч. автономной нервной системы. Интеграция (объединение) деятельности различных отделов автономной Высшим интегративным центром автономной нервной системы являются нервные центры гипоталамуса. Гипоталамус - это образование промежуточного мозга. Задние ядра гипоталамической области являются центрами регуляции симпатичекого отдела автономной нервной системы, передние ядра - высшие интегративные центры парасимпатического отдела автономной нервной системы. Гипоталамические центры могут оказывать свои влияния на нижележащие образования автономной нервной системы через ретикулярную формацию продолговатого мозга, через бульбарные центры: сосудо-двигательный, сердечный, дыхательный и др. Следует отметить, что гипоталамическая область имеет тесные связи с лимбической системой мозга, и в этой связи, данные взаимоотношения позволяют обеспечить взаимосвязь вегетативных механизмов с мотивационноэмоциональными реакциями организма, сомато-вегетативных процессов. Наконец, корковый уровень регуляции вегетативных функций позволяет обеспечить возможность формирования условных вегетативных рефлексов, произвольной регуляции некоторых вегетативных функций.
Лекция №10 ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ. План лекции 1. Общий план строения центральной нервной системы. 2. Структурно-функциональная организация спинного мозга: а) характеристика нейронов, нервных центров и проводящих путей спинного мозга; б) рефлекторная функция спинного мозга; в) проводниковая функция спинного мозга. 1. Общий план строения центральной нервной системы Центральная нервная система состоит из спинного и головного мозга. Головной мозг делят на продолговатый мозг, Варолиев мост, средний мозг, промежуточный мозг, мозжечок и конечный мозг. Конечный мозг состоит из больших полушарий, покрытых корой, белого вещества и базальных ганглиев (см. рис.1). Данная лекция посвящена структурно-функциональной организации спинного и продолговатого мозга. 2. Структурно-функциональная организация спинного мозга Спинной мозг является самым древним в филогенетическом отношении образовании центральной нервной системы. Он состоит из нескольких отделов: шейного, грудного, поясничного, крестцового и копчикового. Каждый организован на основе сегментарного принципа. Каждый сегмент на поперечном сечении спинного мозга состоит из серого вещества (скопление нейронов), белого вещества (проводящие пути спинного мозга), передних и задних корешков. Передние корешки состоят из нервных волокон эфферентных нейронов, а задние корешки из нервных волокон чувствительных нейронов. В соответствии с законом, открытым независимо друг от друга английским физиологом Чарльзом Беллом (1811) и французским физиологом Франсуа Можанди (1822) в составе передних корешков спинного мозга содержатся нервные волокна, покидающие спинной мозг (центробежные волокна), а в составе задних корешков
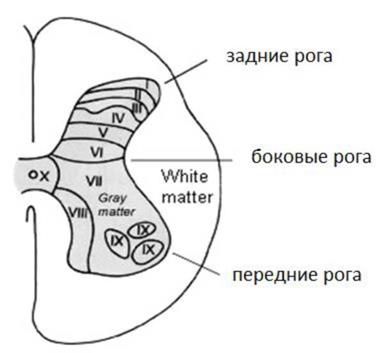
Впоследствии было показано, что афферентных волокон, поступающих в спинной мозг через задние корешки, в несколько раз больше, чем эфферентных, выходящих из спинного мозга. В спинном мозгу различают 31 сегмент. Каждый задний корешок спинного мозга связан не только со своим сегментом, но и с соседними - верхним и нижним через систему межсегментарных связей. Каждый отдел спинного мозга содержит определенное количество сегментов: шейный сегмент содержит 8 сегментов, грудной - 12 сегментов, поясничный - 5 сегментов, крестцовый - 5 сегментов, копчиковый - от 1 до 3 сегментов. а) Характеристика нейронов, нервных центров и проводящих путей спинного мозга В сером веществе спинного мозга содержится более 10 000 000 нервных клеток. Из них 3% составляют ассоциативные, вставочные нейроны (интернейроны), а 97% - исполнительные или эфферентные нейроны. Среди эфферентных нейронов, локализованных в передних рогах спинного мозга, выделяют а1 а2 и у мотонейроны. В скелетных мышцах а1 мотонейроны иннервируют белые или быстрые мышечные волокна, а2 мотонейроны иннервируют красные или медленные мышечные волокна, а у мотонейроны иннервируют периферические сократительные части волокон-рецепторов (ин-трафузальных волокон). Перечисленные выше мотонейроны отличаются не только по тому, какие элементы скелетных мышц они иннервируют, но и по ряду физиологических особенностей. В частности по свойству лабильности. Так а1 мотонейроны способны генерировать потенциалы действий лишь с частотой 30 - 60 имп/сек., а2 мотонейроны с частотой 10 - 20 имп/сек., а у мотонейроны с частотой 150 - 300 имп/сек. Отдельный эфферентный нейрон иннервирует большое количество мышечных волокон, образуя двигательную единицу. В свою очередь скопление эфферентных нейронов иннервирующих одну мышцу образует мотонейронный пул. В боковых рогах спинного мозга грудного, поясничного и крестцового отделов спинного мозга локализуются нейроны автономной нервной системы, для которых характерна постоянная низкочастотная импульсная активность. В соответствии с представлениями Рекседа (1964) нейроны спинного мозга, объединенные по функциональным признакам, в спинном мозге формируют пластины (см. рис. 2).
Рис.2 Функциональная топоргафия нейронов спинного мозга по Рекседу (1964). Обозначения: I - IV - сенсорная зона. Нейроны данных пластин имеют синаптические входы от афферентных нейронов, несущих информацию от экстерорецепторов туловища и конечностей; V - VI - нейроны данных пластин имеют синаптические входы от афферентных нейронов, несущих информацию от проприорецепторов мышц туловища и конечностей. Посредством задних канатиков связаны с сенсомоторной корой больших полушарий. VII - преганглионарные нейроны автономной нервной системы, а также интернейроны, обеспечивающие связь нейронные популяции спинного мозга; VIII - интернейроны, обеспечивающие связь между правой и левой сторонами; IX - эфферентные нейроны спинного мозга; X - клетки глии спинного мозга.
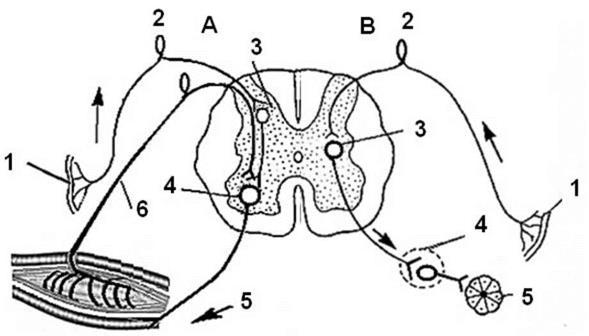
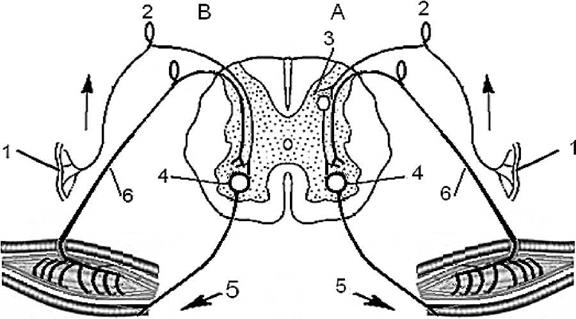
б) Рефлекторная функция спинного мозга Напомню, что рефлексом называют ответную реакцию организма с участием центральной нервной системы при действии раздражителя на чувствительные (сенсорные) рецепторы. Рефлексы спинного мозга разделяют на соматические и рефлексы автономной нервной системы. На рис. 3 показаны спинальные соматический и автономный рефлекс. По критерию времени, в течение которого осуществляется тот или иной рефлекс, спинальные рефлексы можно разделить фазные (бастро- реализующиеся) и тонические (медленнореализующиеся) (см. рис. 5). По критерию числа синапсов в центральной нервной системе у того или иного рефлекса, рефлексы спинного мозга делят на моносинаптические (а) и полисинаптические (б) (см. рис. 4).
Рис. 3 Схема соматического и рефлекса автономной (вегетативной) первной системы спинного мозга. Обозначения А- соматический рефлекс; В - автономный рефлекс; 1- сенсорный рецептор, 2 - чувствительный нейрон, 3 - вставочный нейрон, 4 - эфферентный нейрон, 5 - исполнительный орган, 6 - обратная связь.
Рис.4. Схема рефлексов моносинаптического (В) и полисинаптического (А). 1- сенсорный рецептор, 2 - чувствительный нейрон, 3 - вставочный нейрон, 4 - эфферентный нейрон, 5 - исполнительный орган, 6 - обратная связь.
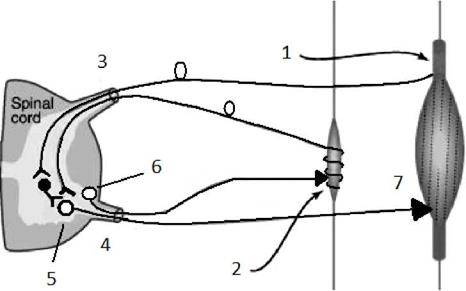
Рис. 5 Схема тонического рефлекса спинного мозга.
Обозначения: 1- сухожильный рецептор Гольджи; 2- мышечное веретено; 3- задние рога спинного мозга; 4-передние рога спинного мозга; 5- 0,2- мотонейрон; 6- у- мотонейрон; 7-мышца. в) Проводниковая функция спинного мозга Проводниковая функция спинного мозга связана с проведением сигналов от чувствительных рецепторов на периферии в спинной мозг, от спинного мозга к вышележащим отделам ЦНС, от вышележащих отделов ЦНС к спинному мозгу и от спинного мозга к исполнительным органам. Афферентная информация в спинной мозг поступает в составе задних корешков, в которых, по последним данным, имеется около 12000 нервных волокон. Все афферентные входы несут в спинной мозг информацию от трех групп сенсорных рецепторов: кожных рецепторов (болевых, температурных, прикосновения, вибрации), проприорецепторов (интрафузальных волокон, сухожильных рецепторов, рецепторов надкостницы и оболочек суставов), рецепторов внутренних органов (висцерорецепторов - механо- осмо- термои хеморепторов). Значение афферентной импульсации, поступающей в спинной мозг, заключается в следующем: - участие в координации деятельности нервных центров; - участие в процессах регуляции функций внутренних органов; - поддержание тонуса ЦНС. Основные проводящие пути спинного мозга и их функции Собственные проводящие пути спинного мозга. Обеспечивают связь между нейронами в пределах одного сегмента спинного мозга или связь между нейронами различных сегментов спинного мозга. Эти пути начинаются от нейронов промежуточной зоны спинного мозга (VII пластина Основные восходящие проводящие пути спинного мозга 1. Проприоцептивные пути, начинающиеся от проприорецепторов мышц (интрфузальных волокон, сухожильных рецепторов). В составе задних корешков афферентные волокна, связанные с проприорепторами мышц поступают в спинной мозг, где контактируют со вставочными нейронами, от которых начинаются тонкий и клиновидный пучки заднего канатика (Голя и Бурдаха), восходящие к продолговатому мозгу, где осуществляется переключение в соответствующих ядрах (нервных центрах). В пределах продолговатого мозга данные проводящие пути осуществляют перекрест (переходят на противоположную сторону) и по медиальному лемнисковому пути направляются к вентромедиальным ядрам зрительного бугра, а затем далее к нейронам IV слоя сенсомоторной коры.
Tractus corticospinalls lateralis Tractus rubrospinalis
Tractus spinotectalis Tractus olivospinalis Tractus reticulospinalis Cornu ventrale Рис. 5 Схема проводящих путей спинного мозга 2. Спиноталамические пути включают латеральный (tractus spinothalamicus) (боковой- lateralis) и вентральный (ventralis) спиноталамические тракты (рис. 5). Латеральный спиноталамический тракт служит для проведения информации о болевых и температурных ощущениях, а вентральный спиноталамический тракт является проводником информации о тактильном чувстве. Аксоны афферентных нейронов, предающих информацию о температурных и болевых ощущениях, направляются в спинной мозг, где формируют синапсы на ассоциативных нейронах VII пластины по Рекседу. Аксоны от этих нейронов поднимаются на несколько сегментов спинного мозга соей стороны, а затем переходят на противоположную сторону, формируя латеральный спиноталамический тракт. Афферентные волокна, несущие информацию от тактильных сенсорных рецепторов, после поступления в спинной мозг в составе задних корешков, заканчиваются на нейронах прмежуточной зоны VII пластины, аксоны которых в пределах данного сегмента через переднюю комиссуру проходят на противоположную сторону и выйдя за пределы серого вещества формируют передний спиноталамический тракт (рис. 5). 3. Вентральный (передний - ventralis) и дорсальный (задний- dorsalis) спино-мозжечковые тракты (tractus spinocerebellaris) (тракты Говерса и Флексига). Ряд афферентных волокон, несущих информацию от проприорецепторов мышц и рецепторов кожи, реагирующих на изменение длины мышц и механического давления, в спинном мозге формируют синапсы на нейронах V и VI пластин, где и начинаются вентральный и дорсальный спиномозжечковые тракты. Основное назначение указанных путей - доставка информации к нервным центрам, участвующим в координации двигательной активности. Кроме указанных проводящих путей, известен ряд других, участвующих в передаче информации от спинного мозга к вышележащим структурам: спиноретикулярные, спинотектальные, спинобульбарные и др. В целом можно отметить, что восходящие пути в спинном мозге занимают в основном задние и заднебоковые отделы белого вещества спинного мозга.
|
|||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 487; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.134.77.195 (0.073 с.) |

 содержатся нервные волокна, (центростремительные волокна).
содержатся нервные волокна, (центростремительные волокна).