Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Способность нервных центров к суммации возбуждения.
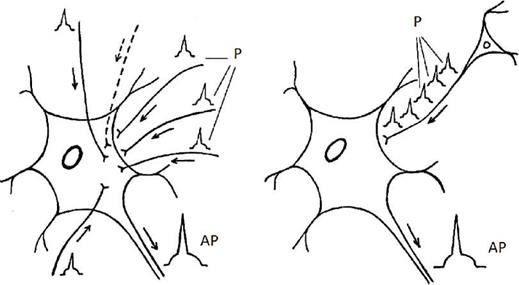
Различают два возможных варианта суммации: временная или последовательная суммация и пространственная суммация. На рисунке 1 А представлены взаимоотношения процессов возбуждения при последовательной (временной) суммации, на рисунке 1 Б - при пространственной суммации. А В
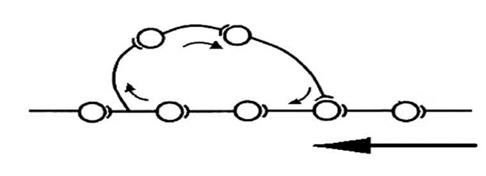
Рис. 1 Взаимоотношения процессов возбуждения при А - пространственной суммации, В - последовательной (временной). Р - афферентная импульсация, АР - Потенциал на аксоне нейрона. При последовательной суммации возбуждение в виде потенциалов действия в нервный центр поступает о одному афферентному входу. Поэтому на постсинаптических мембранах синапсов нервных клеток нервного центра происходит последовательное возникновение постсинаптических потенциалов, которые и суммируются, доводя мембранный потенциал клеток, в отдельные моменты времени, до критического уровня деполяризации и возникновения потенциала действия. При пространственной суммации нервный центр выступает как объемное образование, возбуждение к которому поступает по различным афферентным входам (см. рисунок 2). В этой связи на нейронах нервного центра возникает комплекс постсинаптических потенциалов, которые инициируют возникновение местных токов в соме нейрона и последующую генерацию потенциала действия на аксонном холмике. 4. Высокая чувствительность к химическим соединениям. Это свойство связано со сходством химического строения тех или иных химических соединений и медиаторов. Поэтому указанные соединения могут менять состояние постсинаптических мембран нейронов, входящих в состав нервных центров. 5. Наличие в нервных центрах эффекта посттетанической потенциации. Увеличение эффективности проведения возбуждения через нервный центр в связи с наличием преды-дущего возбуждения. Эффект объясняется потенциацией возбуждения в синапсах, в связи с накоплением в пресинаптических терминалях ионов кальция. 6. Наличие в нервных центрах последействия. Этот феномен объясняется наличием в нервных центрах коллатералей, формирующих так называемые «нейронные ловушки» (см.рис. 2).
Рис. 2 Схема нейронной сети нервного центра, участвующая в формировании эффекта последействия (по Лоренте де Но).
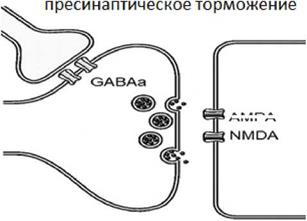
7. Высокая утомляемость нервных центров. Эта особенность связана с большим количеством химических синапсов в нервных центрах, которые характеризуются высокой утомляемостью. 8. Низкая лабильность нервных центров. Это свойство также связано с большим количеством химических синапсов в нервных центрах, которые характеризуются низкой лабильностью. 9. Высокая чувствительность к недостатку кислорода. В нервной ткани преобладают аэробные процессы, поэтому нервные клетки чувствительны к недостатку кислорода. Особенно чувствительны к дефициту кислорода кортикальные нервные центры (нервные центры коры больших полушарий. 10. Способность нервных центров к трансформации ритма. Эта особенность объясняется также наличием большого количества синапсов в нервных центрах, а также особенностями физиологических свойств нервных клеток. 11. Высокая пластичность нервных центров. Способность нервных центров изменять свою специализацию для компенсации функций поврежденных нервных центров. 3. Координация в нервной системе, роль торможения в механизмах координации. Координация в нервной системе - это процесс согласования возбуждения и торможения нервных центров, участвующих в реализации тех или иных рефлексов. Как следует из определения, в механизмах координации принимает участие торможение. Виды торможения в центральной нервной системе. 1. По механизму торможение делят на гиперполяризационное и деполяризационное или первичное и вторичное. 2. По локализации торможение делят на пресинаптическое и постсинаптическое. Пресинаптическое торможение реализуется в нейронных сетях в условиях взаимодействия двух пресинаптических терминалей (см. рис. 3).
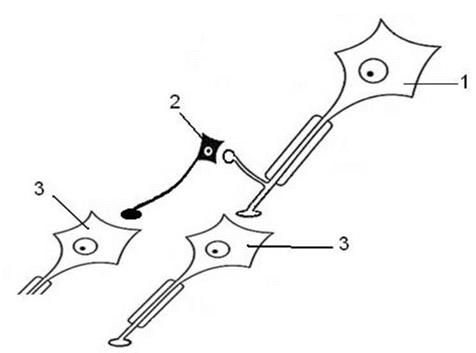
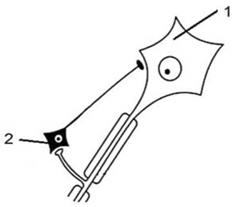
Постсинаптическое торможение реализуется на постсинаптической мембране. По механизму пресинаптическое и постсинаптическое виды торможения могут быть как первичным, так и вторичным. 3. Реципрокное или взаимоисключающее торможение. Нервные центры в нервной системе могут находиться в реципрокных или взаимоисключающих отношениях. Это означает то, что если возбужден один из двух центров, находящихся в реципрокных отношениях, то второй - заторможен. Одним из механизмов таких отношений является механизм реципрокного торможения (см.рис. 4), примером которого является торможение нервных центров мышц-антагонистов, заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей. Если бы возбуждались одновременно центры мышц-сгибателей и мышц- разгибателей, сгибание конечности в суставе было бы невозможно.
Рис. 4. Схема отношений между нейронами ЦНС при реализации реципрокного торможения. Обозначения: 1- возбуждающий (асоциативный) нейрон, 2- тормозный нейрон, 3- возбуждающий (исполнительный) нейрон. 4. Возвратное торможение. Этот вариант торможения впервые описал Б. Реншоу (см. рис. 5) - это такое торможение, при котором тормозные вставочные нейроны (клетки Реншоу) действуют на те же нервные клетки, которые их иннервируют. Примером возвратного постсинаптического торможения может служить торможение в мотонейронах спинного мозга.
Рис. 5 Схема отношений между нейронами ЦНС при реализации возвратного торможения. Обозначения: 1- возбуждающий нейрон, 2 - тормозный нейрон.
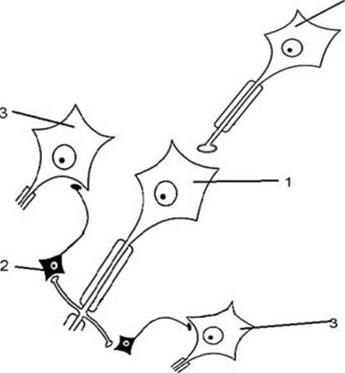
Тормозные нейроны, которые впервые описал американский нейрофизиолог Бэрдсли Реншоу (1911-1948), характеризуются низкой возбудимостью, поэтому их возбуждение возможно только при большой частоте ПД поступающих им на вход. Совершенно очевидно, что в этой ситуации возвратное торможение выполняет, прежде всего, охранительновосстановительную функцию. 5. Латеральное торможение. Данный вид торможения реализуется по следующей схеме (см. рис. 6). Тормозные интернейроны (их тела выделены черным цветом) возбуждаются возвратными коллатералями переключательных сенсорных нейронов: чем сильнее возбужден переключательный нейрон, тем большее торможение соседних переключательных нейронов он вызовет.
Рис. 6 Схема отношений между нейронами в ЦНС при реализации латерального торможения. Обозначения: 1- возбуждающий (переключательный сенсорный) нейрон, 2 - тормозный нейрон, 3 - возбуждающий (исполнительный (переключательный)) нейрон.
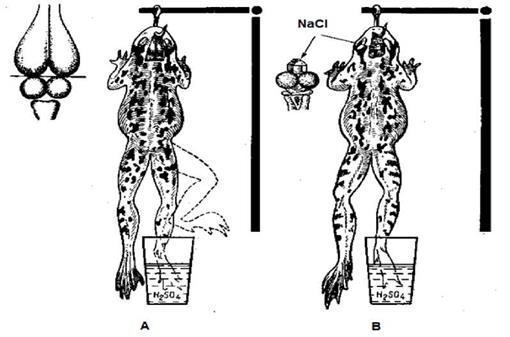
Существует классификация тормозных механизмов на основании того, в каком отделе нервной системы этот механизм реализуется. По этому критерию торможение делят на центральное и периферическое. Периферическое торможение впервые в своих исследованиях в 1845 году открыли немецкие ученые братья Эрнст и Эдуард Вебер. Они показали, что при раздражении периферической ветви блуждающего нерва, иннервирующей сердце, происходит уменьшение частоты сердечных сокращений и уменьшение сердечного выброса крови. Центральное торможение в 1862 году, впервые, было описано в работах И.М. Сеченова. И.М. Сеченов проводил исследования на таламической лягушке. Таламическая лягушка - это лягушка с удаленным конечным мозгом, но сохраненным промежуточным мозгом. Одной из ведущих структур промежуточного мозга является зрительный бугор (thalamus). Поэтому такая лягушка и получила название таламической. На первом этапе у такой лягушки И.М. Сеченов исследовал время рефлекса по Тюрку (защитный рефлекс при воздействии на кожу лапки 1% раствором серной кислоты H2SO4) (рис.7). Затем на зрительный бугор лягушки он помещал кристаллик NaCl и отмечал увеличение времени оборонительного рефлекса. Схему, отражающий механизм центрального торможения можно представить следующим образом: (см. рис. 8).
Рис.7.Опыт проведенный И.М.Сеченовым по выявлению торможения (И.М. Сеченов, 1862). А — двигательный рефлекс на болевой раздражитель; В — раснространснне нервных импульсов от тормозных нейронов ствола мозга к спинному мозгу при наложении кристалла NaCI на область зрительных чертогов и отсутствие двигательного рефлекса на болевой раздражитель
Принципы координации. 1. Принцип реципрокной иннервации. Этот принцип подчеркивает важность механизма реципрокных отношений в механизмах координации деятельности нервных центров. 2. Принцип общего конечного пути или «воронки» Ч. Шерингтона. Этот принцип подчеркивает тот факт, что исполнительных клеток в центральной нервной системе значительно меньше, чем других. Например, в спинном мозге эфферентные клетки составляют всего лишь 3% от всех нейронов. Поэтому в центральной нервной системе отмечается постоянная «борьба» за конечный, исполнительный путь. 3. Принцип доминанты. Этот принцип нашел отражение в работах известного Российского физиолога А.А. Ухтомского в 30 годы ХХ столетия. Термин «доминанта» означает главенствующий, превосходящий. Этот принцип подчеркивает тот факт, что в центральной нервной системе можно обнаружить главенствующие на данный момент времени, доминирующие нервные центры, которые обеспечивают реализацию тех или иных наиболее важных физиологических функций. Доминирующие нервные центры могут возникать как на основе рефлекторных процессов, так и на основе действия гуморальных факторов. Доминирующие нервные центры обладают рядом свойств, которые и обеспечивают им главенствующее положение в ЦНС. К таким свойствам относят: повышенную возбудимость, повышенную способность к суммации возбуждения, выраженную способность вытормаживать активность других нервных центров, инертность возбуждения. 4. Принцип обратной связи. Этот принцип подчеркивает значимость механизмов обратной связи в согласованной деятельности нервных центров (П.К. Анохин, Н.А. Бернштейн и др.).
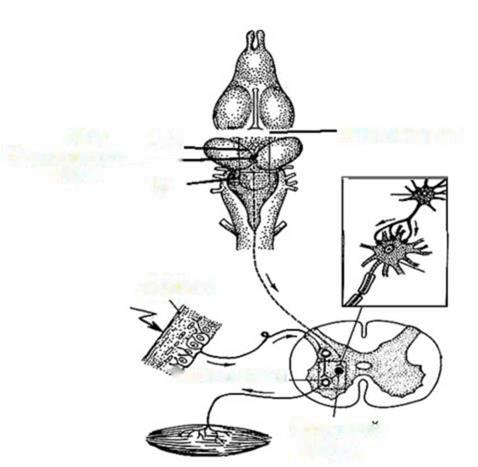
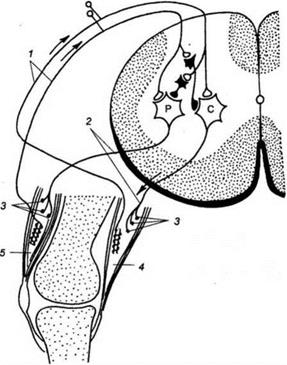
5. Принцип временной связи. Этот принцип подчеркивает роль приобретенных в онтогенезе связей между нерными центрами для согласованной их деятельности. Примером «обогащения» механизмов координации временными связями может служить акт ходьбы у человека, который формируется у детей в первые 1,5 - 2 года жизни на основе врожденного механизма - так называемого рефлекса «шагания». «Рефлекс шагания» - это механизм согласованной активности нервных центров, иннервирующих мышцы антогонисты на конечностях - сгибание одной ноги сопровождается разгибанием другой. Схема реализации «рефлекса шагания» представлена на рис. 8.
Рис. 8. Схема «рефлекса шагания». Обозначения: 1 - афферентая связь от интрафузальный волокон; 2 -аксоны эфферентных нейронов; 4 - musculus flexor, 5 - musculus extensor, P и C - мотонейроны передних рогов спинного мозга. Описанный спинальный механизм «рефлекса шагания» находится под контролем вышележащих отделов ЦНС, в том числе коры головного мозга и используется организмом при формировании акта ходьбы. Лекция №9 ФИЗИОЛОГИЯ АВТОНОМНОЙ (ВЕГЕТАТИВНОЙ) НЕРВНОЙ План лекции. 1. Исторические аспекты развития представлений об автономной (вегетативной) нервной системе. 2. Структурно-функциональная организация автономной нервной системы. Сравнительная характеристика вегетативного и соматического рефлексов. Сравнительная характеристика симпатического и парасимпатического отделов автономной нервной системы. 3. Структурно-функциональная организация метасимпатического отдела автономной нервной системы. 4. Интеграция (объединение) деятельности различных отделов автономной нервной системы в интересах организма. Исторические аспекты развития представлений об автономной Под автономной (вегетативной) нервной системой понимают комплекс центральных и периферических нейронных образований, обеспечивающих согласованную деятельность внутренних органов, с целью приспособления его к изменяющимся условиям окружающей среды. Первые сведения, касающиеся представлений о структуре и функциях автономной нервной системы, связаны с именем известного римского врача Галена (129 - 199 гг. н.э.). Именно Гален дал название «симпатический» (единокровный, обеспечивающий связь) нервному стволу, располагающемуся вдоль позвоночного столба, описал ход и распространение внутри организма блуждающего нерва (Х пара черпно- мозговых нервов). Гален полагал, что симпатические нервы обеспечивают гармонизацию деятельности внутренних органов. Более подробное описание симпатических ганглиев было сделано анатомом из Падуи (Италия) Андреем Везалием в 16 столетии. А. Везалий впервые высказал мысль о том, что симпатические нервы обеспечивают связь внутренних органов с головным мозгом. Значительным этапом формирования представлений об автономной нервной системе были работы известного французского анатома Франсуа Биша (1801), который разделил все функции организма на животные (соматические) и органические. Он полагал, что животная (соматическая) жизнь обеспечивается спинным мозгом, в то время как органическая поддерживается симпатической узелковой (ганглиозной) системой.
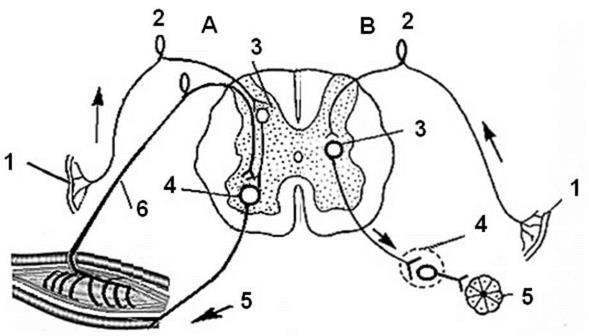
Немецкий анатом И. Рейл (J. Reil) в 1908 году ввел понятие “вегетативная (растительная) нервная система”. Важным этапом развития представлений об автономной нервной системе явились работы английского физиолога Вильяма Гаскела (18471914). Гаскел впервые конкретизировал источники вегетативной нервной системы: черепной отдел, грудопоясничный и крестцовый. Кроме того Гаскелом было сформулировано положение об антагонистической симпатико-парсимпатической иннервации внутренних органов и трофической функции вегетативной нервной системы. Одновременно с В. Гаскеллом, российский физиолог И.П. Павлов (1886) высказал предположение о том, что симпатический и блуждающий нервы оказывают влияние не только на частоту и силу сердечных сокращений, но и изменяют питание (трофику) сердечной мышцы. Во многом близкие к современным, представления о структурно-функциональной организации автономной нервной системы сформировались благодаря работам известного английского нейрофизиолога Джона Ленгли (1852-1925). Благодаря работам Д. Ленгли автономную нервную систему разделили на два самостоятельных отдела симпатический и парасимпатический. Кроме того, он выделил в самостоятельный отдел кишечную систему микро ганглиев (аурбахово и мейснеровсое нервные сплетения тонкой кишки). Последний отдел автономной нервной системы в нашей стране, благодаря работам А.Д. Ноздрачева, называется метасимпатическим отделом. Ограничение во времени не позволяет упомянуть всех ученых, которые внесли свою лепту в развитие представлений об автономной нервной системе, поэтому позвольте перейти к изложению следующей части нашей лекции. Структурно-функциональная организация автономной нервной Автономную нервную систему, как уже отмечалось выше, делят на три отдела: симпатический, парасимпатический и метасимпатический. Симпатический и парасимпатический отделы, также как и соматический, имеют рефлекторную организацию. Вместе с тем, вегетативный рефлекс отличается от соматического (см. рис. 1).
Рис. 1 Схемы рефлекторных дуг соматического и вегетативного рефлексов. Обозначения: А - соматический рефлекс; В - вегетативный рефлекс. 1- рецептор; 2 - афферентный нейрон; 3 - ассоциативный (вставочный) нейрон; 4 - эффекторный нейрон; 5. -эффектор; 6 - обратная связь;
|
|||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 305; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.112.220 (0.038 с.) |