Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Размножение и индивидуальное развитие организмов
Жизнь на Земле сохраняется благодаря тому, что организмы воспроизводят себе подобных. Рост и размножение живых организмов – это результат деления клеток. Без деления невозможно представить себе существование и развитие сложного многоклеточного организма из одной оплодотворенной яйцеклетки. Переход от клеточного уровня к многоклеточному организменному уровню невозможен без рассмотрения механизмов деления клетки. Увеличение числа бактерий связано с бинарным делением их клеток. При этом материнская клетка делится на две дочерние, приблизительно равной величины. В дочерних клетках все компоненты клеточных структур, органеллы и ядро представлены в тех Гаметы образуются в специализированных органах – половых железах. Особый вид деления клеток, в результате которого образуются половые клетки, называется мейозом. При мейозе число хромосом в дочерних клетках уменьшается вдвое. Связано это с тем, что деление клеток происходит без разделения хромосом, поэтому половые клетки получают только половину того числа хромосом, которое содержится в соматических клетках. Таким образом, все соматические клетки организма содержат диплоидный (двойной) набор хромосом, где каждая хромосома имеет парную, гомологическую хромосому. Зрелые половые клетки имеют лишь гаплоидный (одинарный) набор хромосом и соответственно вдвое меньше количество ДНК.
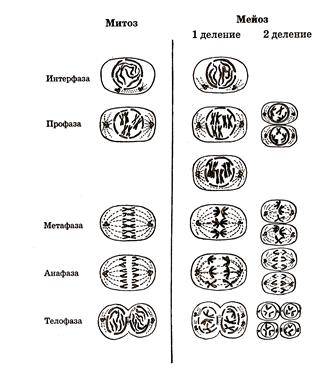
Процесс мейоза состоит из двух последовательных этапа – мейоза I и мейоза II, каждый из которых включает три фазы – профазы (метафазы), анафазы и телофазы (рису-
а) б)
Рисунок 135
В профазе I происходит спирализация хромосом. Гомологичные хромосомы соединя- ются друг с другом по всей длине и скручиваются. Этот процесс соединения гомологичных хромосом носит название конъюгации. Во время конъюгации между некоторыми гомологичными хромосомами происходит обмен участками – генами (кроссинговер), что означает обмен наследственной информацией. В метафазе I гомологичные хромосомы отделяются друг от друга, образуется веретено деления и хромосомы располагаются в плоскости экватора. Затем наступает анафаза I мейоза, и к полюсам клетки отходят хромосомы, каждая из которых состоит из двух хроматид. Следовательно, в дочернюю клетку попадает только одна из каждой пары гомологичных хромосом. Телофаза I длится недолго. Гомологичные хромосомы попадают в разные клетки с гаплоидным набором хромосом. Вслед за процессом мейоза I начинается второе деление – мейоз II, которое сопровож-дается разделением центромер и превращением каждой хроматиды в самостоятельную хромосому (анафаза II). В телофазе II завершается расхождение сестринских хромосом к полюсам, и наступает деление клетки. В результате из двух гаплоидных клеток образуются четыре гаплоидные дочерние клетки. На одной из стадий мейоза (профаза I) происходит взаимный обмен частями между ДНК, содержащейся в хромосомах. Расхождение каждой пары гомологических хромосом определяет закономерности передачи наследственной информации от родителей потом-ству. Из каждой пары двух гомологичных хромосом (материнской и отцовской), входив-ших в хромосомный набор диплоидных организмов, в гаплоидном наборе яйцеклетки или сперматозоиде содержится лишь одна хромосома. Она может быть: 1) отцовской хромосо-мой; 2) материнской хромосомой; 3) отцовской с участками материнской; 4) материнской с участием отцовской. Возникновение большого количества качественно различных половых клеток способствует наследственной изменчивости. В результате слияния двух гаплоидных половых клеток образуется зигота, начальная стадия развития зародыша. При этом зигота диплоидна, поскольку содержит двойной набор хромосом.
Развитие сложного многоклеточного организма из одной оплодотворенной яйцеклетки происходит на основе клеточного деления – митоза. Период жизни клетки от одного деления до следующего называется клеточным циклом. Во время мейоза происходит точная передача наследственной информации дочерним клеткам, поскольку они получают такой же диплоидный набор хромосом, какой был в материнской клетке. Различают следующие фазы митоза: интерфаза, профаза, метафаза, анафаза и телофаза. Ход митоза приведен на рисунке 135б. В интерфазе происходит подготовка клетки к делению. Идет интенсивный синтез белка, АТФ и других органических веществ, необходимых для роста будущих дочерних клеток. Удваиваются все важнейшие структуры клетки. Вдоль исходной хромосомы из химических соединений, находящихся в клетке, происходит синтез ее точной копии, удваивается количество ДНК. Удвоенная хромосома состоит из двух половинок – хроматид, каждая из которых содержит одну молекулу ДНК. В профазе происходит спирализация и вследствие этого утолщение хромосом, состоящих из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредотачиваются по всей клетке. В цитоплазме к концу профазы центроли отходят к полюсам и образуется веретено деления. В метафазе происходит дальнейшая спирализация хромосом. Их центромеры располагаются по экватору. К ним прикрепляются нити веретена деления. В анафазе центромеры делятся, сестринские хромосомы отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки. В телофазе цитоплазма делится, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. Так из исходной клетки (материнской) образуются две новые – дочерние с диплоид-ным набором хромосом. Таким образом, индивидуальное развитие многоклеточных организмов (онтогенез) и их размножение обеспечиваются особыми клеточными механиз-мами – митозом и мейозом. Эмбриональное развитие большинства высших организмов протекает по единому плану и включает три основных этапа: дробление, гаструляцию, органогенез. Развитие многоклеточных животных начинается с митотических делений зиготы. На начальном этапе деления оплодотворенной яйцеклетки образуются многоклеточные бластомеры. Бластомеры не растут и при каждом последующем делении дочерних клеток уменьшаются вдвое. Поэтому процесс деления яйцеклетки в этом случае называется дроблением. Заключительная фаза процесса дробления зиготы сопровождается образованием бластулы – однослойного зародыша. У многих животных она имеет форму однослойного бластодермического пузырька, заполненного жидкостью.
После дробления начинается процесс гаструляции (от греч. gaster – желудок), который сопровождается перемещением клеточного материала с поверхности бластулы во внутрь на места будущих органов. В результате гаструляции образуется гаструм, состоящий из обособленных зародышевых листков: наружного (эктодермы), среднего (мезодермы) и внутреннего (эндодермы). Зародышевые листки являются зачатками органов, приведенных в таблице 14. Таблица 14
Следует отметить, что точного соответствия между тремя зародышевыми листками и органами животных не существует. В образовании некоторых органов участвуют два, а иногда даже три листка. Так, эндодерма образует выстилку кишечника, а поддержи-вающие его ткани возникают из мезодермы. Развитие многоклеточных организмов из одной единственной оплодотворенной яйцеклетки выдвинуло перед молекулярной биологией сложнейшую проблему. Суть ее заключается в том, что все клетки организма идентичны по генотипу, но различны по фенотипу. Они существуют как специализированные элементы многоклеточных высших животных. Согласно современным представлениям молекулярной биологии строгое соответствие между генотипом и фенотипом является основным ее принципом. Чтобы не подвергать сомнению центральное положение современной биологии необходимо было ответить на ряд вопросов. Каким образом происходит специализация клеток? Какой механизм координирует развитие различных клеток, организует построение из них органов и тканей? Как происходит развитие целостного организма? Эти и другие проблемы, связанные с функционированием живой материи, получили разрешение в рамках концепции дифференциальной экспрессии генов. Биохимическая специализация клеток обеспечивается дифференцированной актив-ностью (экспрессией) их генов. Гены могут как включаться, так и выключаться. Диф-ференцированная экспрессия генов означает их неодинаковое проявление в фенотипе. Кроме того, были обнаружены такие участки ДНК, которые управляют деятельностью генов. Поэтому, когда говорят о дифференциальной экспрессии генов, то имеют в виду весь комплекс процессов, обеспечивающих существование и рост организмов.
Механизм возникновения дифференциации деятельности генов до конца не изучен, но выявлены факторы, регулирующие активность генов. Обнаружено три типа факторов регуляции генов: генные, белковые и физико-химический фактор воздействия на клетки извне. К генным факторам управления экспрессией генов относятся промоторы, энхансеры, транспозоны и метилированные ДНК. Промотор – участок ДНК, узнаваемый РНК-полимеразой и инициирующий транс-крипцию. Без распознавания промотора генные механизмы не запускаются. Энхансеры (от анг. enhance – усиливать), – регуляторные участки ДНК, которые активируют транс-крипцию, находясь на расстоянии в несколько тысяч нуклеотидных пар. Наглядный пример – нервная клетка, ядро которой, расположенное в спинном мозге, содержит генетическую информацию для синтеза особых ферментов, которые нужны в окончаниях отростков, иногда удаленных от ядра на десятки сантиметров. Транспозоны – фрагменты ДНК, способные отделяться и внедряться в нее, ускоряя эволюцию геномов. Метилирование ДНК состоит в переводе цитозина в 5-метилцитозин под действием ферментов. Установлено, что метилирование ДНК регулирует активность генов и формирует генную память: раз возникнув, она сохраняется в ряду клеточных поколений. Таким образом, ДНК содержит ряд факторов, регулирующих активность генов, что непосредственно отражается на фенотипе через генотип. Транскрипция генов существенно зависит от так называемых белков-регуляторов, механизм функционирования которых рассмотрен ранее. Важную роль в механизме экспрессии генов в клетках играют сигналы из внешней среды, в том числе от соседних клеток. Таким образом, проблема – ДНК одна и та же, а клетки организма разные – разрешается на основе концепции дифференциальной экспрессии генов. Современная биология дает не только феноменологическое описание всего многообразия живых организмов, но и объясняет его на молекулярно-генетическом уровне.
|
||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 104; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.104.29 (0.014 с.) |