Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Химический синапс, его ультраструктура. Механизм передачи сигнала в химическом синапсе. Механизм возникновения постсинаптического потенциала.
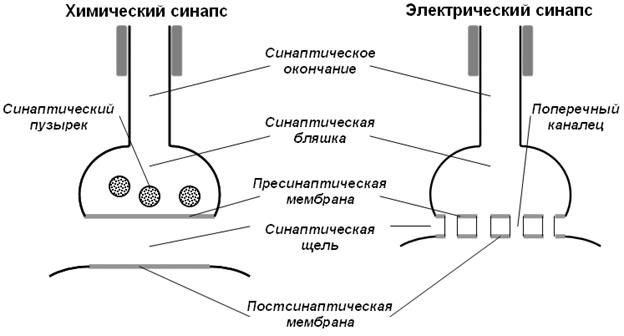
Ультраструктура химического синапса. Все синапсы имеют общий план строения (рис. 1). Конечная часть аксона (синаптическое окончание), подходя к иннервируемой клетке, теряет миелиновую оболочку и образует на конце небольшое утолщение (синаптическую бляшку). Ту часть мембраны аксона, которая контактирует с иннервируемой клеткой, называют пресинаптической мембраной. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках). Синаптическая щель – узкое пространство между пресинаптической мембраной и мембраной иннервируемой клетки, которое является непосредственным продолжением межклеточного пространства. Постсинаптическая мембрана – участок мембраны иннервируемой клетки, контактирующий с пресинаптической мембраной через синаптическую щель. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Особенности ультраструктуры химического синапса (см. рис. 1): • широкая (20–50 нм) синаптическая щель; Постсинаптические рецепторы – это трансмембранные белки, чья наружная часть узнаёт и связывает молекулы медиаторов. Связь рецептора с медиатором специфична. Рецепторы также связаны с ионными каналами в мембране. Существует два способа управления этими каналами и, соответственно, два типа рецепторов: а) Ионотропный (прямая синаптическая передача). При этом виде управления рецептор и ионный канал представляют собой единую молекулу. Если к рецептору присоединяется медиатор, то изменяется конформация молекулы и образуется пора, через которую проходят ионы. Пример – никотиновый АХ-рецептор. б) Метаботропный (непрямая синаптическая передача). В этом случае рецептор не связан с каналом напрямую (они разделены), поэтому моменты присоединения медиатора и открытия канала разделены несколькими промежуточными этапами. Когда образуется комплекс медиатор-рецептор, активируется специфический мембранный белок, называемыый G-белком. Одна молекула медиатора может активировать много молекул G-белка (происходит усиление). В одних нейронах каждая активировання молекула G-белка может открывать ионный канал, а к других нейронах – активировать внутри клетки синтез специфических молекул – вторичных посредников. Последние могут запускать в клетке различные биохимические реакции (например, синтез определённых белков). В этом случае не возникает постсинаптический потенциал. Некоторые вторичные посредники (напр. NO и CO) могут диффундировать в соседние нейроны и там оказывать своё действие (в т.ч. и на пресинаптические нейроны).
Т.о. ионотропное управление является очень быстрым и занимает 1-2мс. При метаботропном управлении ответ появляется значительно позже – через несколько секунд или минут, но при этом изменения в клетке сохраняются дольше, могут усиливатья, давая более долговременный эффект. Вещества, которые оказывают на постсинаптические рецепторы такое же действие, как и медиатор, называются агонистами, а вещества, которые связываются с рецептором и блокируют их, называются антагонистами. Эти термины обычно применяются в фармакологических веществах. Нарушение синаптических контактов может привести к тяжелейшим последствиям для человека. Антагонисты могут использоваться в лечебных целях. Механизм проведения возбуждения в химическом синапсе путем освобождения трансмиттеров. Механизм открытия каналов у ионотропных рецепторов Серия потенциалов действия пробегает по аксону, достигает нервного окончания и деполяризует пресинаптическую зону. Во время этой деполяризации в нервном окончании возникает не только входящий ток натрия, как это происходит в мембране по всей длине аксона. Мембрана окончания аксона имеет потенциалуправляемые кальциевые каналы, через которые во время деполяризации, вызванной пришедшим потенциалом действия, ионы кальция проникают в синаптическое окончание, так как там исходно очень низкая концентрация кальция. Одновременно ионы кальция дополнительно выходят из эндоплазматического ретикулума. В синаптическом окончании в зоне пресинаптической мембраны имеется большое количество синаптических пузырьков (везикул). Их мембраны состоят из бислоя фосфолипидов и белков. Эти везикулы заполнены жидкостью, содержащей химическое вещество – трансмиттер, благодаря которому осуществляется синаптическая передача.Трансмиттер переносит возбуждение от пресинаптической мембраны на постсинаптическую. Одним из таких веществ является ацетилхолин. Синапс, в котором в качестве трансмиттера из везикул выбрасывается ацетилхолин, называется холинергическим.
Если потенциал действия достиг пресинаптической области и в пресинаптическом окончании концентрация ионов кальция достигла необходимого уровня, происходят 2 процесса. Во-первых, на везикулах, связанных с пресинаптической мембраной, кальций связывается с протеином, входящим в состав их мембраны. Это приводит к тому, что мембранные везикулы раскрываются, происходит экзоцитоз трансмиттера в синаптическую щель. Повышенный уровень ионов кальция в пресинапсическом окончании активирует кальций-кальмодулинзависимую протеинкиназу 2. В пресинаптическом окончании этот фермент фосфорилирует синапсин (протеин, с помощью которого везикула фиксируется большей частью на цитоскете). После этого нагруженные трансмиттером везикулы освобождаются от цитоскелета и перемещаются на пресинаптическую мембрану для осуществления дальнейшего цикла. Таким образом, главную роль для процесса выброса трансмиттера по типу экзоцитоза играет не деполяризация окончания, а входящий ток кальция. Ионы кальция при этом служат не для дополнительной деполяризации, а вторичным посредником, запускающим механизм слияния везикул. Механизм открытия каналов у метаботропных рецепторов В противоположность синапсам (например никотиновым), в которых трансмиттер открывает ионный канал, существуют другие рецепторные белки, не являющиеся ионными каналами. Примером может служить холинергический синапс мускаринового типа. Название синапс приобрел по действию агониста – яда мухомора мускарина. В этом синапсе Ach-рецептором является белок. Этот белок обладает большим химическим сходством со светочувствительным пигментом родопсином, α- и β- адренергическими и другими рецепторами. Ионные каналы, необходимые для возникновения ВПСП, открываются там только благодаря обменным процессам. Поэтому их функция включает процессы метаболизма, а эти рецепторы называют метаботропными. Процесс передачи возбуждения в этом синапсе происходит следующим образом. Как только медиатор связывается с рецептором, G-белок, имеющий три субъединицы, образует с рецептором комплекс. В этом родопсин, мускариновый рецептор, и все другие рецепторы, связанные с G-белками, похожи друг на друга. ГДФ, связанный с G-белком, заменяется на ГТФ. При этом образуется активированный G-белок, состоящий из ГТФ и α-субъединицы, который и открывает калиевый ионный канал.
Механизм возникновения постсинаптического потенциала. Взаимодействуя с рецептором, молекулы ацетилхолина открывают неспецифические ионные каналы в постсинаптической мембране клетки так, что повышается их способность к проводимости для одновалентных катионов. Какие катионы проходят через каналы, зависит от электрохимических градиентов. Работа данных каналов ведет к базовому входящему току положительных ионов, следовательно к деполяризации постсинаптической мембраны (ВПСП). Так как участвующие ионные токи зависят от разности равновесного потенциала и потенциала мембраны, то при уменьшенном потенциале покоя мембраны ток ионов натрия ослабевает, а ток ионов калия увеличивается, поэтому амплитуда ВПСП уменьшается.
Ионные токи, участвующие в возникновении ВПСП, ведут себя иначе нежели токи натрия и калия во время генерации потенциала действия. Причина в том, что в этом механизме принимают участие другие ионные каналы с другими свойствами. При потенциале действия активируются потенциалуправляемые каналы, и с увеличивающейся деполяризацией открываются следующие каналы, так что процесс деполяризации усиливает сам себя, а проводимость лигандуправляемых каналов зависит от количества молекул трансмиттера, связавшихся с молекулами рецептора и от числа открытых ионных каналов. Прежде всего, в зоне синапса локально образовавшийся ВПСП пассивно электротонически распространяется по всей постсинаптической мембране клетки. Это распространение не подчиняется закону «все или ничего». Если большое число синапсов возбуждается одновременно или почти одновременно, то возникает явление суммации, которое проявляется в возникновении ВПСП большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны определенного порогового значения (10 мВ или больше), то на аксональном холмике нервной клетки молниеносно открываются потенциалуправляемые натриевые каналы и она генерирует потенциал действия, проводящийся вдоль ее аксона.
|
||||||||||
|
|
Последнее изменение этой страницы: 2021-01-08; просмотров: 383; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.188.11 (0.011 с.) |