Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нервное волокно: функциональное значение отдельных структурных элементов, классификация нервных волокон. Механизм проведения возбуждения по миелинизированным и немиелинизированным волокнам.Стр 1 из 13Следующая ⇒
Сравнительная характеристика электрических и химических синапсов. Их физиологические свойства, чувствительность к внешним регуляторным воздействиям.
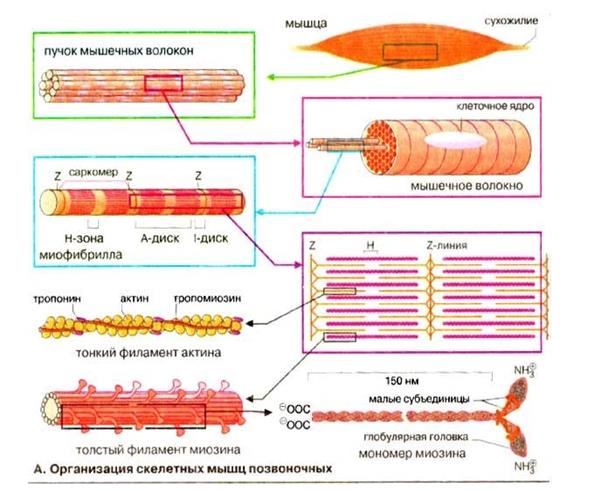
Скелетная мышца: функциональное значение отдельных структурных элементов мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы и двигательного аппарата организма, классификация двигательных единиц Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл, которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Толстые филаменты состоят из белка миозина, тонкие-из актина, тропопнина и тропомиозина. Миозиновые филламенты образованы повторяющимися молекулами белка миозина. Каждая молекула миозина имеет головку и хвост. Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик. Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++
Светлая(изотропная)полоса-I-диск, расположена между краями А-дисков двух соседних саркомеров и состоит из тех участков, которые не перекрываются толстыми филаментами. Z-пластинка делит I-диск пополам.
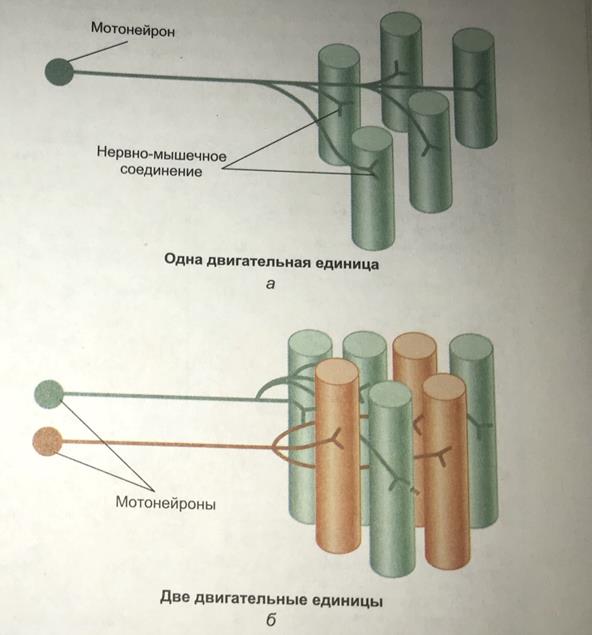
Волокна скелетных мышц иннервируются аксонами нервных клеток--мотонейронами(или соматическими эфферентными нейронами). Тела этих клеток расположены в спинном мозге или в стволе мозга. Мотонейрон и мышечные волокна, которые он иннервирует, составляют двигательную единицу.
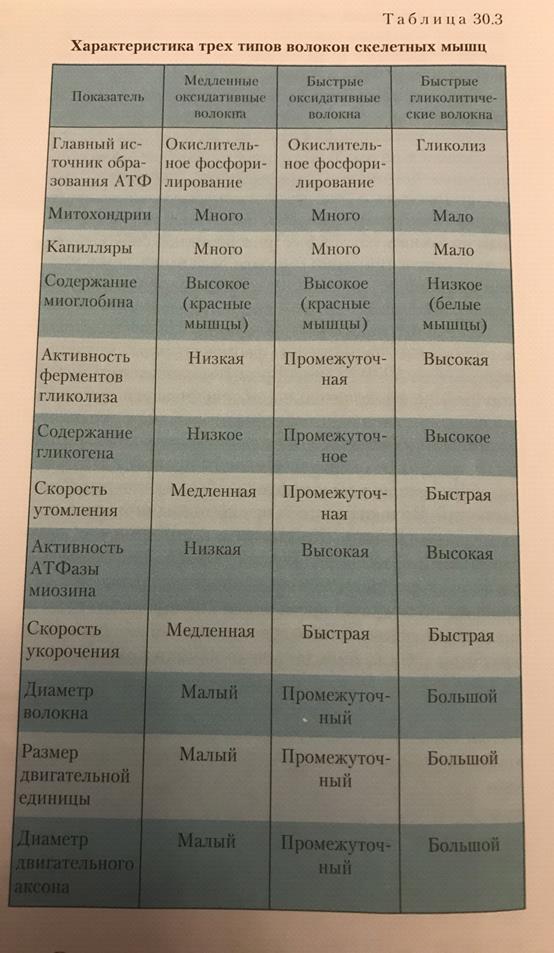
Область плазматической мембраны, лежащая непосредственно под нервным окончанием, обладает особыми свойствами и называется двигательной концевой пластинкой. Структура, состоящая из нервного окнчания и двигательной концевой пластинки, --это нервно-мышечное окончание(нервно-мышечный синапс). Типы двигательных единиц: -Имеют относительно большой диаметр -Отличаются светлым цветом, поскольку в них снижено количество миоглобина -Окружены небольшим количеством капилляров -Имеют относительно небольшое количество митохондрий -Характеризуются высоким содержанием гликогена
Получают АТФ за счет гликолиза. Из-за небольшого количества капилляров и миоглобина получают мало кислорода. Незначительно количество митохондрий в цитозоле также определяет небольшое потребление кислорода. Высокое содержание гликогена обуславливает достаточное количество глюкозы для гликолиза. Посему такие волокна называют гликолитическими. Красные мышечные волокна:
-Диаметр красных мышечных волокон примерно в 2 раза меньше диаметра белых -Темно-красный цвет из-за высокого содержания миоглобина -Окружены большим количеством капилляров -Имеют множество митохондрий -Низкое содержание гликогена Для синтеза АТФ используют цикл Кребса и окислительное фосфорилирование. Небольшой диаметр капилляров облегчает диффузию кислорода. Из-за небольшого содержания гликогена для энергетического обеспечения красных волокон используются жирные кислоты. Они распадаются до ацетил КоА и включатся в цикл Кребса. Посему они называются оксидативными.
1. Медленные оксидативные(тип1)-низкая активность миозиновой АТФазы и высокая окислительная способность. 2.Быстрые оксидативные(тип2а)-высокая активность миозиновой АТФазы и высокая гликолитическая способность. 3.Быстрые гликолитические волокна(тип2б)-высокая активность миозиновой АТФазы и высокая гликолитическая способность.
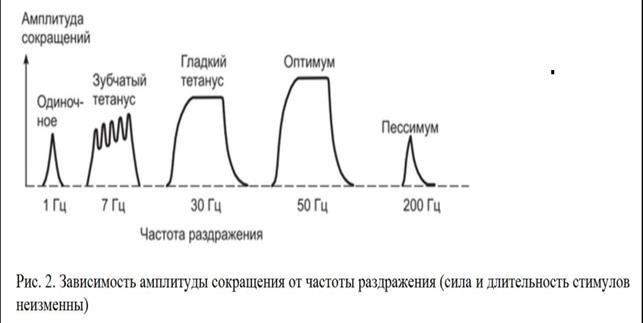
Тетаническое сокращение изолированной мышцы: понятие о тетанусе, механизм, факторы, влияющие на величину тетануса, оптимум и пессимум частоты раздражения. Механизм тетануса в естественных условиях. Работа скелетной мышцы, ее утомление Тетанусом называют сокращение мышцы, возникающее в результате суммирования сокращений ее моторных единиц, вызванных поступлением к ним множества нервных импульсов от моторных нейронов, иннервирующих данную мышцу. Суммирование усилий, развиваемых при сокращении волокон множества двигательных единиц, способствует увеличению силы тетанического сокращения мышцы и влияет на длительность сокращения. Различают зубчатый и гладкий тетанус. Для наблюдения в эксперименте зубчатого тетануса мышцы ее стимулируют импульсами электрического тока с такой частотой, чтобы каждый последующий стимул наносился после фазы укорочения, но еще до окончания расслабления. Гладкое тетаническое сокращение развивается при более частых раздражениях, когда последующие воздействия наносятся во время развития укорочения мышцы. Например, если фаза укорочения мышцы составляет 50 мс, фаза расслабления — 60 мс, то для получения зубчатого тетануса необходимо раздражать эту мышцу с частотой 9-19 Гц, для получения гладкого — с частотой не менее 20 Гц. Для демонстрации различных видов тетануса обычно используют графическую регистрацию на кимографе сокращений изолированной икроножной мышцы лягушки. Пример такой кимограммы представлен на рис. 2. Если сравнивать амплитуды и усилия, развиваемые при различных режимах сокращения мышцы, то они при одиночном сокращении минимальны, увеличиваются при зубчатом тетанусе и становятся максимальными при гладком тетаническом сокращении. Одной из причин такого возрастания амплитуды и силы сокращения является то, что увеличение частоты генерации ПД на мембране мышечных волокон сопровождается увеличением выхода и накоплением в саркоплазме мышечных волокон ионов Са2+, способствующего большей эффективности взаимодействия между сократительными белками. При постепенном увеличении частоты раздражения нарастание силы и амплитуды сокращения мышцы идет лишь до определенного предела — оптимума ответной реакции. Частоту раздражения, вызывающую наибольший ответ мышцы, называют оптимальной. Дальнейшее увеличение частоты раздражения сопровождается уменьшением амплитуды и силы сокращения. Это явление называют пессимумом ответной реакции, а частоты раздражения, превышающие оптимальную величину — пессимальными.
В естественных условиях частота и режим посылки моторными нейронами нервных импульсов к мышце обеспечивают асинхронное вовлечение в процесс сокращения большего или меньшего (в зависимости от числа активных мотонейронов) количества двигательных единиц мышцы и суммацию их сокращений. Сокращение целостной мышцы в организме но своему характеру близко к гладкотеганическому. Для характеристики функциональной активности мышц оценивают показатели их тонуса и сокращения. Тонусом мышцы называют состояние длительного непрерывного напряжения, вызванное попеременным асинхронным сокращением ее моторных единиц. При этом видимое укорочение мышцы может отсутствовать из-за того, что в процесс сокращения вовлекаются не все, а лишь те двигательные единицы, свойства которых наилучшим образом приспособлены к поддержанию тонуса мышцы и силы их асинхронного сокращения недостаточно для укорочения мышцы. Сокращения таких единиц при переходе от расслабления к напряжению или при изменении степени напряжения называют тоническими. Кратковременные сокращения, сопровождаемые изменением силы и длины мышцы, называют физическими.
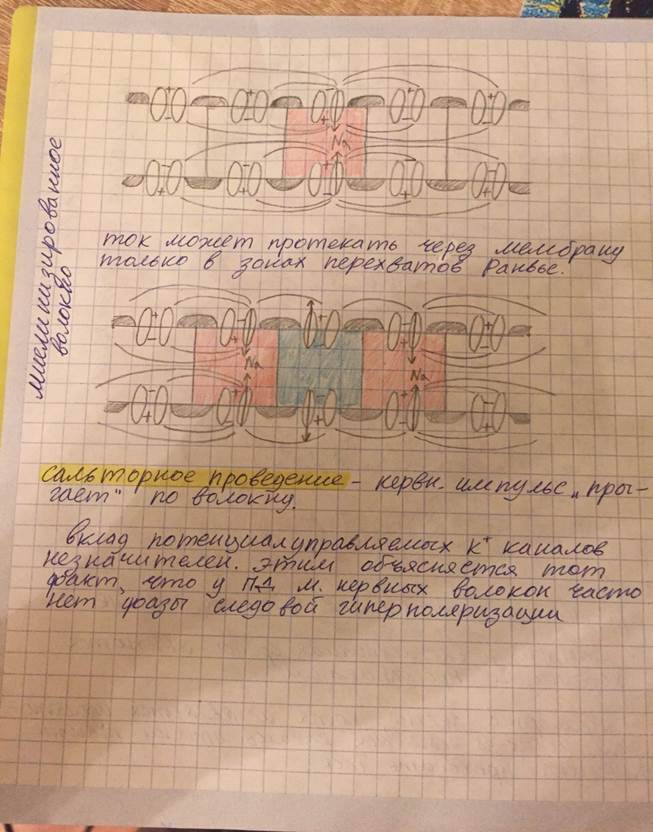
Нервное волокно: функциональное значение отдельных структурных элементов, классификация нервных волокон. Механизм проведения возбуждения по миелинизированным и немиелинизированным волокнам. Функциональное значение структурных компонентов нейрона По строению оболочек различают миелиновые и безмиелиновые нервные волокна (ещё называют мякотные и безмякотные) Строение нервного волокна
Механизм проведения нервного импульса по безмиелиновым волокнам.
* безмиелиновое нервное волокно имеет Na+ каналы на протяжении всей своей длины. Механизм проведения нервного импульса по миелиновым волокнам
|
||||||||
|
|
Последнее изменение этой страницы: 2021-01-08; просмотров: 889; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.140.198.173 (0.014 с.) |