Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Метаболизм клеток соединительной ткани
Фибробласты и их разновидности в специализированных тканях, такие как остеобласты, хондробласты, кератобласты участвуют в синтезе основных компонентов внеклеточного вещества. Этим определяются особенности их метаболизма. В фибробластах синтезируются коллагены, эластин, кератин и ряд других белков внеклеточного вещества. В этих клетках высокая скорость синтеза и распада нуклеиновых кислот, главным образом, матричных РНК кодирующих аминокислотную последовательность белков внеклеточного матрикса, тРНК и рРНК. Высокая интенсивность белкового синтеза в этих клетках обеспечивается повышенным содержанием (по сравнению с другими клетками) ряда аминокислот, таких как глицин, пролин, a- аланин, глютаминовой и аспарагиновой кислот. Содержание этих аминокислот в разы выше, чем в клетках других тканей. Для синтеза белков необходима энергия, которая обеспечивается аэробным окислением глюкозы. Часть глюкозы, поступающая в фибробласты расходуется на синтез глюкуроновой, идуроновой и галактуроновой кислот необходимых для синтеза гликозоаминогликанов, таких как гиалуроновая кислота, хондроитинсульфаты, гепарансульфаты и др. Другая часть глюкозы трансформируется в аминосахара и их ацетилированные производные, которые также входят в состав гликозоаминогликанов. В свою очередь гликозоаминогликаны входят в состав протеогликанов, основного компонента внеклеточного матрикса, которые также образуются в фибробластах. Фибробласты не только синтезируют компоненты внеклеточного матрикса, но и активно участвуют в его обновлении и регуляции состава. Такое обновление достигается выделением фибробластами ферментов участвующих в деградации составных частей этого матрикса. Внеклеточный матрикс, образуемый фибробластами обеспечивает контакты между клетками, придает механическую прочность, изолирует одни ткани от других, обеспечивает обмен водой и электролитами между клетками и кровью и выполняет также ряд других функций. Одним из компонентов внеклеточного матрикса является коллаген, структура, функция и синтез которого рассматривается ниже.
Коллаген
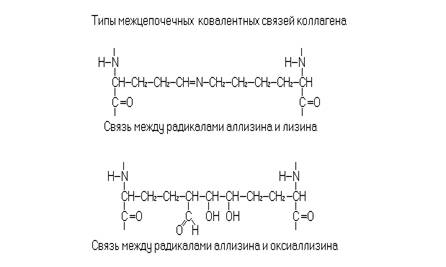
Коллаген один из основных белков соединительной ткани. Белок располагается во внеклеточном веществе, где образует нерастворимые нити, погруженные в матрикс из протеогликанов. Коллагеновые волокна построены из фибрилл. Ориентация фибрилл различается в разных тканях. Коллагеновые волокна связок, сухожилий и кожи ориентированы параллельно, а в заживающих ранах – хаотично. Структурной единицей фибрилл является молекула тропоколлагена. Тропоколлагены выделенные из разных структур отличаются по своему аминокислотному составу, но имеют одинаковую структурную организацию. Типичная молекула тропоколлагена состоит из трех полипептидных цепей с суммарной массой 300000 дальтон. Отдельная полипептидная цепь содержит около 1000 аминокислотных остатков и образует левозакрученную спираль, содержащую 3 остатка на 1 виток. В свою очередь отдельные субъединицы объединяются с образованием правозакрученной спирали, где переплетение отдельных цепей образует структуру наподобие кабеля. Отдельные цепи взаимодействуют между собой за счет водородных связей между пептидными группами соседних цепей. Кроме того, структура стабилизируется ковалентными сшивками между радикалами лизина, оксилизина, аллизина и оксиаллизина. Характерной чертой построения протомеров тропоколлагена является наличие чередующихся последовательностей – Гли - Про - Х -. Часть радикалов пролина и лизина гидроксилирована. В большинстве коллагенов присутствуют 4 – оксипролин и 5 – оксилизин, но в некоторых коллагенах встречается и 3 - оксипролин. Коллагеновые фибриллы образуются молекулами тропоколлагена соединенными конец к концу и бок о бок. Начало молекул в соседних цепях смещено на четверть длины. Боковые взаимодействия молекул тропоколлагена осуществляются за счет не ковалентных и ковалентных взаимодействий.
Синтез коллагена в фибробластах и остеобластах идет на полисомах. Синтезируются все три полипептидные цепи. Все синтезируемые цепи длиннее, чем те которые присутствуют в тропоколлагене. Так например a- цепи содержат дополнительные фрагменты на N- конце молекулярной массой 20000 и C конце 35000 дальтон. Гидроксилирование пролина и лизина осуществляется пролиноксидазой и лизиноксидазой соответственно ещё на стадии элонгации. Вновь синтезированные цепи объединяются в структуру проколлагена. Дополнительные фрагменты на N и C концах образуют глобулярные структуры. Проколлаген секретируется из клетки аппаратом Гольджи. Процессинг проколлагена осуществляется внеклеточно путем гидролиза соответствующих пептидных связей N и C концевыми проколлагеновыми пептидазами.
Помимо гидроксилирования молекулы проколлагена подвергаются гликозилированию с участием нуклеотид – сахар – гликозилтрансфераз. Идет присоединение остатков галактозы или галактозилглюкозы к ОН группе 5 – гидроксилизина. При образовании молекул тропоколлагена и их объединения в фибриллы между a- цепями формируются поперечные ковалентные связи. Сначала лизилоксидаза окисляет концевую -NH2 группу лизина или гидроксилизина до альдегидной группы –СНО. При этом образуются аллизин и оксиаллизин соответственно. Остатки аллизина взаимодействуют с остатками лизина и оксилизина соседних цепей, образуя ковалентную связь.
При конденсации остатков аллизина и оксиаллизина образуется синдезин. Синдезин выделен из зубов человека. В костях, в том числе и зубах коллаген индуцирует минерализацию. Формирование минеральной кристаллической решетки начинается в зоне находящейся между коллагеновыми фибриллами. В процессе формирования кости кристаллы образуются сначала в зоне коллагеновых волокон. Затем они сами в свою очередь становятся центрами нуклеации для отложения кристаллов гидроксиапатита в пространстве между коллагеновыми волокнами. Формирование кости происходит только в непосредственной близости от остеобластов. В зоне кальцификации происходит деградация комплексов белок – полисахарид (протеогликанов) при участии лизосомальных протеиназ клеток кости. Так, по-видимому, протекает минерализация в дентине и цементе зуба. С возрастом состав и структура коллагенов организма меняется. Эти изменения выражаются в снижении растворимости коллагена, увеличении прочности волокон, снижении эластичности и способности к набуханию, изменению содержания оксипролина, в характере гликозилирования.
БИОХИМИЯ КОСТНОЙ ТКАНИ
Кости человека состоят из органического матрикса, кристаллов фосфорно – кальциевых солей 45-50% и клеток кости – остеобластов, остеоцитов, остеокластов. Остеобласты – клетки мезенхимного происхождения, вырабатывают органические составные части остеоида и участвуют в процессах его минерализации. В период активного функционирования эти клетки называются активными остеобластами. После окружения себя минеральным матриксом клетки теряют свою метаболическую активность, меняют форму, теряют способность к размножению, такие клетки называются остеоцитами и преобладают в зрелой кости. Остеоциты окружены полостью (лакуной) и взаимодействуют между собой через многочисленные отростки проходящими в узких костных канальцах. С помощью отростков клетки обмениваются между собой низкомолекулярными метаболитами и ионами через щелевые контакты. Остеобласты оказавшиеся на поверхности кости образуют слой покоящихся остеобластов. Эти клетки образуют тонкий 0,1-0,5 мкм органический матрикс, который не минерализуется. Вырабатываемый ими органический матрикс отделяет их от кости и называется эндостальной мембраной. Такая форма остеоида останавливает рост кости в ширину и защищает её поверхность от «несанкционированной» атаки клетками разрушающими кость. Остеоциты и покоящиеся остеобласты сохраняют способность реагировать на гормональные и другие регуляторные воздействия.
Остеокласты не вырабатывают костную ткань, а участвуют в её резорбции. Эти клетки образуются путем слияния клеток крови моноцитов. Остеокласты содержат десятки ядер, а их размер составляет 20 – 100 мкм. Клетки отличаются высокой активностью карбоангидразы, кислой фосфатазы. В мембранах клеток имеется Н+ - АТФаза и рецепторы к кальцитонину и витронектину (определяет адгезию к окружающему матриксу). Многочисленные лизосомы накапливают и выделяют в среду множество кислых гидролаз, участвующих в расщеплении макромолекул остеоида. Органические компоненты костной ткани представлены, в основном, белками. Протеогликанов и воды в кости гораздо меньше, чем в хряще. Волокнистые структуры костного матрикса на 90% процентов состоят из коллагена 1 типа. Из минорных компонентов присутствуют коллагены типов 5 и 12, которые входят в состав волокон или ассоциированы с ними. Коллаген типа 5 способен связываться с протеогликанами и рядом белков. Считается, что он контролирует конечный диаметр коллагеновых фибрилл. Коллаген 12 типа кроме аналогичных свойств имеет несколько RGD – центров связывания с клетками и, следовательно, участвует в единении клеточных элементов и межклеточных структур. В небольших количествах имеется и коллаген 6 типа. Это белок также имеет RGD – центры и участвует в прикреплении клеток к коллагенам 1 и 5, а также взаимодействия их с неколлагеновыми молекулами. Содержание гиалуроновой кислоты в костях невелико, равно как и количество протеогликанов (декорина и бигликана). Неколлагеновые белки представлены гликопротеинами, которые составляют около 10% органического матрикса. Среди этих белков преобладают сиалопротеины (остеопонтин и костный сиалопротеин 2), остеонектин, костный и матриксный Gla – протеины (белки, которые содержат остатки g- карбоксиглутаминовой кислоты). Остеопонтин и остеонектин содержатся также в мозге, почке, печени и плаценте. Сиалопротеин 2 и остеокальцин характерны только для кости и дентина. Кроме перечисленных белков в кости встречаются и белки плазмы.
Минеральная составляющая кости представлена, главным образом, кристаллами гидроксиапатита, структурированных во внеклеточном матриксе. В целом упаковка частиц в кристалле гидроксиапатита представляет многослойную гексагональную структуру, где с каждым ионом фосфата соседствует 12 ионов кальция и ОН-, из которых 6 лежат в одном слое с фосфатом, а по 3 в соседних слоях, расположенных выше и ниже. Идеальный кристалл гидроксиапатита имеет форму гексагональной призмы, размеры которой варьируют в разных твердых тканях. Расстояние между кристаллами гидроксиапатитов составляет 2,5 нм. Каждый кристалл окружен гидратной оболочкой толщиной 1 нм. Часть ионов кристаллической решетки кристалла гидроксиапатитов может быть замещена другими ионами близкими по свойствам (изоморфное замещение). В местностях с высоким содержанием стронция, накопление его в костях может привести к патологическим переломам. Снижение концентрации кальция и фосфата в жидкостях тела приводит к усилению реакций изоморфного замещения. Напротив, повышенное поступление Са2+ в организм способствует вытеснению из ионной решетки антагонистов кальция, обеспечивая восстановление правильной кристаллической решетки. В целом скорость и масштаб изоморфного замещения зависят, в основном, от ионного радиуса, концентрации и длительности воздействия замещающего иона. Очень большое значение имеют условия микроокружения, в том числе физико-химические параметры, включая рН среды. В частности, достаточно высокий уровень бикарбонатов способствует образованию карбонапатита путем замещения фосфат-иона карбонатом.
Са10(РО4)6(ОН)2 + 3 Н+ + 3 НСО3- ® Са10(РО4)4(СО3)3(ОН)2 + 2 Н3РО4 Среди всех апатитов твердых тканей карбонапатиты могут составлять до 20%, уступая в этом отношении гидроксиапатитам. На хлорапатиты приходится около 4%. Снижение рН среды способствует замещению Са2+ в гидроксиапатите протонами или ионами гидроксония.
Са10(РО4)6(ОН)2 + 2 Н+ ® Са9(Н+)2(РО4)6(ОН)2 + Са2+
Са10(РО4)6(ОН)2 + Н3О+ ® Са9(Н3О+) (РО4)6(ОН)2 + Са2+
Высокие концентрации Н+ вызывают кислотное разрушение кристаллов гидроксиапатита.
Са10(РО4)6(ОН)2 + 8 Н+ ® 10 Са2+ + 6 НРО42- + 2 Н2О
Помимо кристаллов гидроксиапатита и его аналогов минеральная фаза содержит и аморфные соли. В основном это кальция гидрокарбонат [Са(НСО3)2], кальция гидрофосфат [СаНРО4 × 2 Н2О], пентагидрат восьми кальциевого фосфата [Са8Н2(РО4)6 × 5 Н2О]. Аморфные фосфаты преобладают в раннем возрасте и выполняют роль легко мобилизуемого резерва ионов кальция и фосфата, используемого при построении кристаллических структур.
|
||||||||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 237; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.31.240 (0.017 с.) |