
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Последовательность событий, приводящих к изменению скорости транскрипцииСодержание книги
Поиск на нашем сайте Гормон проходит через двойной липидный слой клеточной мембраны. В цитозоле или ядре гормон взаимодействует с рецептором. Комплекс гормон-рецептор проходит в ядро и присоединяется к регуляторной нуклеотидной последовательности ДНК - энхансеру (рис. 4.18) или сайленсеру. Доступность промотора для РНК-полимеразы увеличивается при взаимодействии с энхансером или уменьшается при взаимодействии с сайленсером. Соответственно увеличивается или уменьшается скорость транскрипции определенных структурных генов. Зрелые мРНК выходят из ядра. Увеличивается или уменьшается скорость трансляции определенных белков. Изменяется количество белков, которые влияют на метаболизм и функциональное состояние клетки. В каждой клетке существуют рецепторы, включенные в состав разных сигнал-трансдукторных систем, преобразующих все внешние сигналы во внутриклеточные. Число рецепторов для конкретного первичного мессенджера может варьировать в пределах от 500 до более 100 000 на клетку. Они располагаются на мембране отдаленно друг от друга либо сосредоточены в определенных ее участках.
141.Роль основных компонентов (липидов, белков, углеводов) в структурной организации и функционировании мембран. Все клетки и внутриклеточные органеллы окружены мембранами, которые играют важную роль в их структурной организации и функционировании. Основные принципы построения всех мембран одинаковы. Однако плазматическая мембрана, а также мембраны эндоплазматического ретикулума, аппарата Гольджи, митохондрий и ядра имеют существенные структурные особенности, они уникальны по своему составу и по характеру выполняемых функций. Мембраны: • отделяют клетки от окружающей среды и делят ее на компартменты (отсеки); • регулируют транспорт веществ в клетки и органеллы и в обратном направлении; • обеспечивают специфику межклеточных контактов; • воспринимают сигналы из внешней среды. Согласованное функционирование мембранных систем, включающих рецепторы, ферменты, транспортные системы, помогает поддерживать гомеостаз клетки и быстро реагировать на изменения состояния внешней среды путем регуляции метаболизма внутри клеток. Биологические мембраны построены из липидов и белков, связанных друг с другом с помощью нековалентных взаимодействий. Основу мембраны составляет двойной липидный слой, в состав которого включены белковые молекулы (рис. 4.1). Липидный бислой образован двумя рядами амфифильных молекул, гидрофобные «хвосты» которых спрятаны внутрь, а гидрофильные группы - полярные «головки» обращены наружу и контактируют с водной средой. 1. Липиды мембран. В состав липидов мембран входят как насыщенные, так и ненасыщенные жирные кислоты. Ненасыщенные жирные кислоты встречаются в два раза чаще чем насыщенные, что определяет текучесть мембран и конформационную лабильность мембранных белков. В мембранах присутствуют липиды трех главных типов - фосфолипиды, гликолипиды и холестерол (рис. 4.2 - 4.4). Чаще всего встречаются глицерофосфолипиды - производные фосфатидной кислоты.
Фосфатидная кислота - это диацилглицеролфосфат. R1, R2 - радикалы жирных кислот (гидрофобные «хвосты»). Со вторым углеродным атомом глицерола связан остаток полиненасыщенной жирной кислоты. Полярной «головкой» является остаток фосфорной кислоты и присоединенная к нему гидрофильная группа серина, холина, этаноламина или инозитола Существуют также липиды - производные аминоспирта сфингозина. Аминоспирт сфингозин при ацилировании, т.е. присоединении жирной кислоты к NH2-группе, превращается в церамид. Церамиды различаются по остатку жирной кислоты. С ОН-группой церамида могут быть связаны разные полярные группы. В зависимости от строения полярной «головки» эти производные разделены на две группы - фосфолипиды и гликолипиды. Строение полярной группы сфингофосфолипидов (сфингомиелинов) сходно с глицерофосфолипидами. Много сфингомиелинов содержится в составе миелиновых оболочек нервных волокон. Гликолипиды представляют собой углеводные производные церамида. В зависимости от строения углеводной составляющей различают цереброзиды и ганглиозиды. Холестерол содержится в мембранах всех животных клеток, он придает мембранам жесткость и снижает их жидкостность (текучесть). Молекула холестерола располагается в гидрофобной зоне мембраны параллельно гидрофобным «хвостам» молекул фосфо- и гликолипидов. Гидроксильная группа холестерола, как и гидрофильные «головки» фосфо- и гликолипидов,
Церамид - ацилированный сфингозин (R1 - радикал жирной кислоты). К фосфолипидам относятся сфингомиелины, у которых полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Гидрофильной группой (полярной «головкой») гликолипидов является углеводный остаток. Цереброзиды содержат моноили олигосахаридный остаток линейного строения. В состав ганглиозидов входит разветвленный олигосахарид, одним из мономерных звеньев которого является НАНК - N-ацетилнейраминовая кислота обращена к водной фазе. Молярное соотношение холестерола и других липидов в мембранах равно 0,3-0,9. Самое высокое значение имеет эта величина для цитоплазматической мембраны. Увеличение содержания холестерола в мембранах уменьшает подвижность цепей жирных кислот, что влияет на конформационную лабильность мембранных белков и снижает возможность их латеральной диффузии. При повышении текучести мембран, вызванном действием на них липофильных веществ или перекисным окислением липидов, доля холестерола в мембранах возрастает.
Молекула холестерола состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи. Полярной «головкой» является ОН-группа у 3-го углеродного атома молекулы холестерола. Для сравнения на рисунке представлено схематическое изображение фосфолипида мембран. Полярная головка этих молекул значительно больше и имеет заряд Липидный состав мембран различен, содержание того или другого липида, по-видимому, определяется разнообразием функций, которые выполняют эти молекулы в мембранах. Главные функции липидов мембран состоят в том, что они: • формируют липидный бислой - структурную основу мембран; • обеспечивают необходимую для функционирования мембранных белков среду; • участвуют в регуляции активности ферментов; • служат «якорем» для поверхностных белков; • участвуют в передаче гормональных сигналов. Изменение структуры липидного бислоя может привести к нарушению функций мембран. 2. Белки мембран. Белки мембран различаются по своему положению в мембране (рис. 4.5). Мембранные белки, контактирующие с гидрофобной областью липидного бислоя, должны быть амфифильными, т.е. иметь неполярный домен. Амфифильность достигается благодаря тому, что: • аминокислотные остатки, контактирующие с липидным бислоем, в основном неполярны; • многие мембранные белки ковалентно связаны с остатками жирных кислот (ацилированы). Ацильные остатки жирных кислот, присоединенные к белку, обеспечивают его «заякоревание» в мембране и возможность латеральной диффузии. Кроме того, белки мембран подвергаются таким посттрансляционным модификациям, как гликозилирование и фосфорилирование. Гликозилирование наружной поверхности интегральных белков защищает их от повреждения протеазами межклеточного пространства.
1, 2 - интегральные (трансмембранные) белки; 3, 4, 5, 6 - поверхностные белки. В интегральных белках часть полипептидной цепи погружена в липидный слой. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных «головок», обогащены гидрофильными аминокислотными остатками. Поверхностные белки разными способами прикрепляются к мембране: 3 - связанные с интегральными белками; 4 - присоединенные к полярным «головкам» липидного слоя; 5 - «заякоренные» в мембране с помощью короткого гидрофобного концевого домена; 6 - «заякоренные» в мембране с помощью ковалентно связанного ацильного остатка Наружный и внутренний слои одной и той же мембраны различаются по составу липидов и белков. Эта особенность в строении мембран называется трансмембранней асимметрией. Белки мембран могут участвовать в: • избирательном транспорте веществ в клетку и из клетки; • передаче гормональных сигналов; • образовании «окаймленных ямок», участвующих в эндоцитозе и экзоцитозе; • иммунологических реакциях; • качестве ферментов в превращениях веществ; • организации межклеточных контактов, обеспечивающих образование тканей и органов. 142.Механизмы транспорта веществ через мембраны. 1. Пассивный транспорт может осуществляться следующими способами (рис. 4.6, 4.7):
К пассивному транспорту относится диффузия ионов по белковым каналам, например диффузия Н+, Са2+, N+, К+. Функционирование большинства каналов регулируется специфическими лигандами или изменением трансмембранного потенциала.
ИФ3 (инозитол-1,4,5-трифосфат) образуется при гидролизе мембранного липида ФИФ2 (фосфатидилинозитол-4,5-бисфосфата) под действием фермента фосфолипазы С. ИФ3 связывается специфическими центрами протомеров Са2+- канала мембраны эндоплазматического ретикулума. Изменяется конформация белка и канал открывается - Са2+ поступает в цитозоль клетки по градиенту концентрации 2. Активный транспорт. Первично-активный транспорт происходит против градиента концентрации с затратой энергии АТФ при участии транспортных АТФаз, например Na+, К+-АТФаза, Н+-АТФаза, Са2+-АТФаза (рис. 4.8). Н+-АТФазы функционируют как протонные насосы, с помощью которых создается кислая среда в лизосомах клетки. С помощью Са2+-АТФазы цитоплазматической мембраны и мембраны эндоплазматического ретикулума поддерживается низкая концентрация кальция в цитозоле клетки и создается внутриклеточное депо Са2+ в митохондриях и эндоплазматическом ретикулуме. Вторично-активный транспорт происходит за счет градиента концентрации одного из переносимых веществ (рис. 4.9), который создается чаще всего Na+, К+-АТФазой, функционирующей с затратой АТФ. Присоединение в активный центр белка-переносчика вещества, концентрация которого выше, изменяет его конформацию и увеличивает сродство к соединению, которое проходит в клетку против градиента концентрации. Вторично-активный транспорт бывает двух типов: активный симпорт и антипорт.
|
||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 154; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.73 (0.009 с.) |

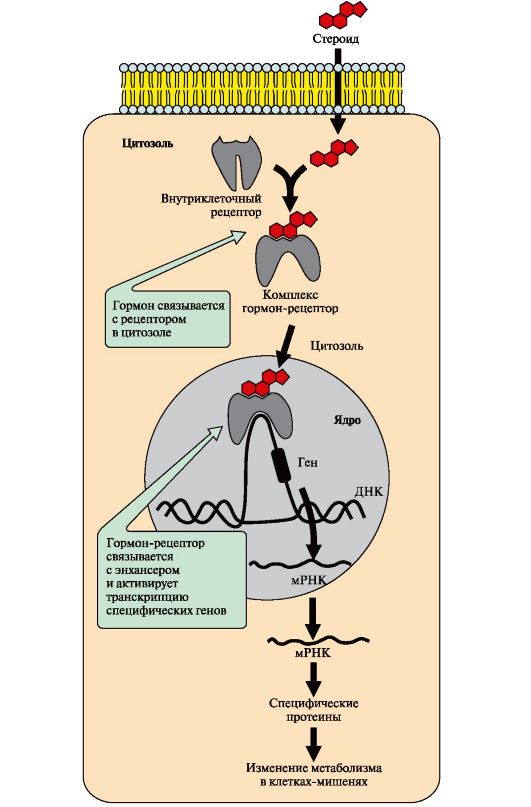 Рис. 4.18. Передача сигнала на внутриклеточные рецепторы
Рис. 4.18. Передача сигнала на внутриклеточные рецепторы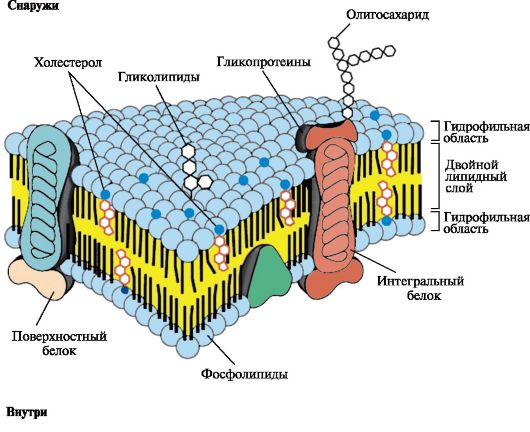 Рис. 4.1. Поперечный разрез плазматической мембраны
Рис. 4.1. Поперечный разрез плазматической мембраны Рис. 4.2. Глицерофосфолипиды.
Рис. 4.2. Глицерофосфолипиды.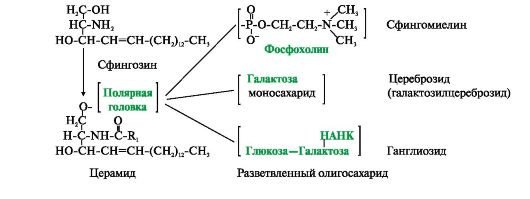 Рис. 4.3. Производные аминоспирта сфингозина.
Рис. 4.3. Производные аминоспирта сфингозина. Рис. 4.4. Положение в мембране фосфолипидов и холестерола.
Рис. 4.4. Положение в мембране фосфолипидов и холестерола. Рис. 4.5. Белки мембран:
Рис. 4.5. Белки мембран: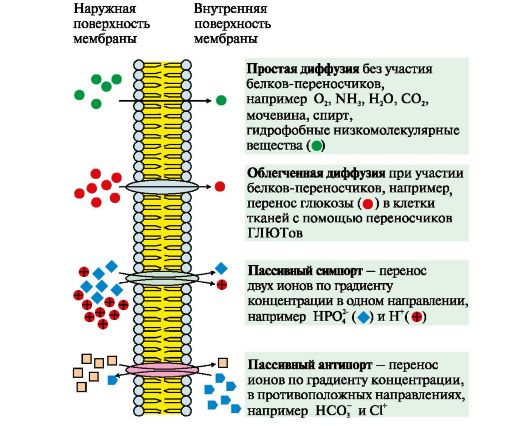 Рис. 4.6. Механизмы переноса веществ через мембраны по градиенту концентрации
Рис. 4.6. Механизмы переноса веществ через мембраны по градиенту концентрации Рис. 4.7. Са2+-канал мембраны эндоплазматического ретикулума, регулируемый инози- тол-1,4,5-трифосфатом (ИФ3).
Рис. 4.7. Са2+-канал мембраны эндоплазматического ретикулума, регулируемый инози- тол-1,4,5-трифосфатом (ИФ3). Рис. 4.8. Механизм функционирования Са2+-АТФазы
Рис. 4.8. Механизм функционирования Са2+-АТФазы Рис. 4.9. Вторично-активный транспорт
Рис. 4.9. Вторично-активный транспорт


