Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизм мышечного сокращения
Рассмотрим, к чему сводятся представления о механизме попеременного сокращения и расслабления мышц. В настоящее время принято считать, что биохимический цикл мышечного сокращения состоит из 5 стадий (рис. 20.8): 1) миозиновая «головка» может гидролизовать АТФ до АДФ и Н3РО4 (Pi), но не обеспечивает освобождения продуктов гидролиза. Поэтому данный процесс носит скорее стехиометрический, чем каталитический, характер (см. рис. 20.8, а); 2) содержащая АДФ и Н3РО4 миозиновая «головка» может свободно вращаться под большим углом и (при достижении нужного положения) связываться с F-актином, образуя с осью фибриллы угол около 90° (см. рис. 22.8, б); 3) это взаимодействие обеспечивает высвобождение АДФ и Н3РО4 из актин-миозинового комплекса. Актомиозиновая связь имеет наименьшую энергию при величине угла 45°, поэтому изменяется угол миозинас осью фибриллы с 90° на 45° (примерно) и происходит продвижение актина (на 10–15 нм) в направлении центра саркомера (см. рис. 20.8, в); 4) новая молекула АТФ связывается с комплексом миозин–F-актин (см. рис. 20.8, г);
Рис. 20.8. Биохимический цикл мышечного сокращения. Объяснение в тексте. 5) комплекс миозин–АТФ обладает низким сродством к актину, и поэтому происходит отделение миозиновой (АТФ) «головки» от F-актина. Последняя стадия и есть собственно расслабление, которое отчетливо зависит от связывания АТФ с актин-миозиновым комплексом (см. рис. 20.8, д). Затем цикл возобновляется. Регуляция сокращения и расслабления мышц. Сокращение любых мышц происходит по общему механизму, описанному ранее. Мышечные волокна разных органов могут обладать различными молекулярными механизмами регуляции сокращения и расслабления, однако всегда ключевая регулятор-ная роль принадлежит ионам Са2+. Установлено, что миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться в его присутствии лишь при наличии в среде определенныхконцентраций ионов кальция. Наибольшая сократительная активность наблюдается при концентрации ионовСа2+ около 10–6–10–5 М. При понижении концентрации до 10–7 М или ниже мышечные волокна теряют способность к укорочению и развитию напряжения в присутствии АТФ. По современным представлениям, в покоящейся мышце (в миофибрил-лах и межфибриллярном пространстве) концентрация ионов Са2+ поддерживается ниже пороговой величины в результате связывания их структурами (трубочками и пузырьками) саркоплазматической сети и так называемой Т-системой при участии особого Са2+-связывающего белка, получившего название кальсеквестрина, входящего в состав этих структур.
Связывание ионов Са2+ разветвленной сетью трубочек и цистерн сарко-плазматической сети не является простой адсорбцией. Это активный физиологический процесс, который осуществляется за счет энергии, освобождающейся при расщеплении АТФ Са2+-зависимой АТФазой саркоплазматической сети. При этом наблюдается весьма своеобразная картина: скорость выкачивания ионов Са2+ из межфибриллярного пространства стимулируется этими же ионами. В целом такой механизм получил название «кальциевая помпа» по аналогии с хорошо известным в физиологии натриевым насосом. Возможность пребывания живой мышцы в расслабленном состоянии при наличии в ней достаточно высокойконцентрации АТФ объясняется снижением в результате действия кальциевой помпы концентрации ионовСа2+ в среде, окружающей миофибриллы, ниже того предела, при котором еще возможны проявление АТФазной активности и сократимость акто-миозиновых структур волокна. Быстрое сокращение мышечного волокна при его раздражении от нерва (или электрическим током) является результатом внезапного изменения проницаемости мембран и как следствие выхода из цистерн и трубочек саркоплазматической сети и Т-системы некоторого количества ионов Са2+ в саркоплазму. Как отмечалось, «чувствительность» актомиозиновой системы к ионам Са2+ (т.е. потеря актомиозиномспособности расщеплять АТФ и сокращаться в присутствии АТФ при снижении концентрации ионов Са2+ до 10–7 М) обусловлена присутствием в контрактильной системе (на нитях F-акти-на) белка тропонина, связанного с тропомиозином. В тропонин-тропомио-зиновом комплексе ионы Са2+ связываются именно с тропонином. В молекуле тропонина при этом происходят конформационные изменения, которые, по-видимому, приводят к сдвигу всего тропонин-тропомиози-нового стержня и деблокировке активных центровактина, способных взаимодействовать с миозином с образованием сократительного комплекса и активной Mg2+-АТФазы.
В продвижении актиновых нитей вдоль миозиновых, по данным Э. Хаксли, важную роль играют временно замыкающиеся между нитями поперечные мостики, которые являются «головками» миозиновых молекул. Итак, чем большее число мостиков прикреплено в данный момент к акти-новым нитям, тем больше сила мышечного сокращения. Наконец, если возбуждение прекращается, содержание ионов Са2+ в саркоплазме снижается (кальциевая помпа), то циклы прикрепление–освобождение прекращаются, т.е. «головки» миозиновых нитей перестают прикрепляться к актиновым нитям. В присутствии АТФ мышца расслабляется и ее длина достигает исходной. Если прекращается поступление АТФ (аноксия, отравление дыхательными ядами или смерть), то мышца переходит в состояние окоченения. Почти все поперечные мостики толстых (миозиновых) нитей присоединены при этом к тонким актиновым нитям, следствием чего и является полная неподвижность мышцы. Мышечные белки А.Я. Данилевский впервые разделил экстрагируемые из мышц белки на 3 класса: растворимые в воде, экстрагируемые 8–12 % раствором хлорида аммония и белки, извлекаемые разбавленными растворамикислот и щелочей. В настоящее время белки мышечной ткани делят на три основные группы: саркоплазматические, миофибриллярные и белки стромы. На долю первых приходится около 35%, вторых – 45% и третьих – 20% от всего количества мышечного белка. Эти группы белков резко отличаются друг от друга по растворимости в воде и солевых средах с различной ионной силой. Белки, входящие в состав саркоплазмы, относятся к протеинам, растворимым в солевых средах с низкой ионной силой. Принятое ранее подразделение саркоплазматических белков на миоген, глобулин X, миоальбумин и белки-пигменты в значительной мере утратило смысл, поскольку существование глобулина X и миогена как индивидуальных белков в настоящее время отрицается. Установлено, что глобулин X представляет собой смесь различных белковых веществ со свойствами глобулинов. Термин «миоген» также является собирательным понятием. В частности, в состав белков группы миогена входит ряд протеинов, наделенных ферментативной активностью: например, ферменты гликолиза. К числу саркоплазмати-ческихбелков относятся также дыхательный пигмент миоглобин и разнообразные белки-ферменты, локализованные главным образом в митохондриях и катализирующие процессы тканевого дыхания,окислительного фосфорилирования, а также многие стороны азотистого и липидного обмена. Недавно была открыта группа саркоплазматических белков – пар-вальбумины, которые способны связывать ионы Са2+. Их физиологическая роль остается еще неясной. К группе миофибриллярных белков относятся миозин, актин и актомио-зин – белки, растворимые в солевых средах с высокой ионной силой, и так называемые регуляторные белки: тропомиозин, тропонин, α- и β-актинин, образующие в мышце с актомиозином единый комплекс. Перечисленные миофибриллярныебелки тесно связаны с сократительной функцией мышц.
Рис. 20.3. Строение молекулы миозина. Объяснение в тексте. Миозин составляет 50–55% от сухой массы миофибрилл. Представление о миозине как о главном белкемиофибрилл сложилось в результате работ А.Я. Данилевского, О. Фюрта, Э. Вебера и ряда других исследователей. Однако всеобщее внимание к миозину было привлечено лишь после опубликования работ В.А. Энгельгардта и М.Н. Любимовой (1939– 1942). В этих работах впервые было показано, что миозинобладает АТФазной активностью, т.е. способностью катализировать расщепление АТФ на АДФ и Н3РО4. Химическая энергия АТФ, освобождающаяся в ходе данной ферментативной реакции, превращается в механическую энергию сокращающейся мышцы. Молекулярная масса миозина скелетных мышц около 500000 (для миозина кролика 470000). Молекула миозина (рис. 20.3) имеет сильно вытянутую форму, длину 150 нм. Она может быть расщеплена без разрыва ковалентных связей на субъединицы: две тяжелые полипептидные цепи с мол. массой 205000–210000 и несколько коротких легких цепей, мол. масса которых около 20000. Тяжелые цепи образуют длинную закрученную α-спираль («хвост» молекулы), конец каждой тяжелой цепи совместно с легкими цепями создает глобулу («головка» молекулы), способную соединяться сактином. Эти «головки» выдаются из основного стержня молекулы. Легкие цепи, находящиеся в «головке» миозиновой молекулы и принимающие участие в проявлении АТФазной активности миозина, гетерогенны по своему составу. Количество легких цепей в молекуле миозина у различных видов животных и в разных типах мышц неодинаково.
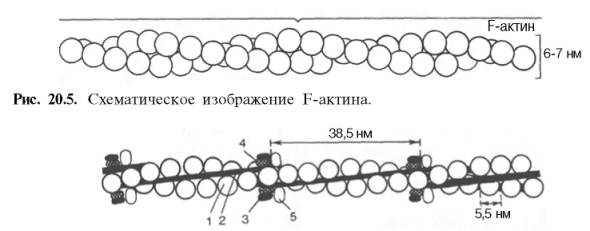
Кратковременная обработка трипсином расщепляет молекулу миозина на два фрагмента. Из хвостового участка (С-концевой участок молекулы) образуется легкий меромиозин (ЛММ) - фрагмент длиной 90 нм, а из остальной части, включающей «головки»,- тяжелый меромиозин (ТММ). ЛММ, подобно миозину, образует нити, однако он не обладает АТФазной активностью и не связывает актин. ТММ катализирует гидролиз АТФ и связывает актин. ТММ можно расщепить далее путем более длительной обработки трипсином илипапаином, в результате чего получается один S2-фрагмент длиной 40 нм с мол. массой 62000 и два S1-фрагмента с мол. массой 110000, представляющие собой «головки» миозина. Толстые нити (толстые миофиламенты) в саркомере надо понимать как образование, полученное путем соединения большого числа определенным образом ориентированных в пространстве молекул миозина(рис. 20.4). Актин, составляющий 20% от сухой массы миофибрилл, был открыт Ф. Штраубом в 1942 г. Известны две формы актина: глобулярный актин (G-актин) и фибриллярный актин (F-актин). Молекула G-актина с мол. массой 42000 состоит из одной полипептидной цепочки (глобула), в обра-
Рис. 20.4. Строение толстого миозинового филамента. зовании которой принимают участие 374 аминокислотных остатка. При повышении ионной силы до физиологического уровня G-актин полиме-ризуется в F-актин (фибриллярная форма). На электронныхмикрофотографиях волокна F-актина выглядят как две нити бус, закрученных одна вокруг другой (рис. 20.5). Актомиозин образуется при соединении миозина с F-актином. Ак-томиозин, как естественный, так и искусственный, т.е. полученный путем соединения in vitro высокоочищенных препаратов миозина и F-актина, обладает АТФазной активностью, которая отличается от таковой миозина, АТФазная активность миозиназначительно возрастает в присутствии стехиометрических количеств F-актина. Фермент актомиозинактивируется ионами Mg2+и ингибируется этилендиаминтетраацетатом (ЭДТА) и высокой концентрациейАТФ, тогда как миозиновая АТФаза ингибируется ионами Mg2+, активируется ЭДТА и не ингибируется высокой концентрацией АТФ. Оптимальные значения рН для обоих ферментов также различны. Как отмечалось, кроме рассмотренных основных белков, в миофибрил-лах содержатся также тропомиозин, тропонин и некоторые другие ре-гуляторные белки. Тропомиозин был открыт К. Бейли в 1946 г. Молекула тропомиозина состоит из двух α-спиралей и имеет вид стержня длиной 40 нм; его мол. масса 65000. На долю тропомиозина приходится около 4–7% всех белковмиофибрилл. Тропонин – глобулярный белок, открытый С. Эбаси в 1963 г.; его мол. масса 80000. В скелетных мышцах взрослых животных и человека тропонин (Тн) составляет лишь около 2% от всех миофибриллярных белков. В его состав входят три субъединицы (Тн-I, Тн-С, Тн-Т). Тн-I (ингибирующий) может ингибировать АТФазнуюактивность, ТН-С (кальцийсвязывающий) обладает значительным сродством к ионам кальция, Тн-Т (тропомиозин-связывающий) обеспечивает связь с тропомиозином. Тропонин, соединяясь с тропомиозином, образует комплекс, названный нативным тропомиози-ном. Этот комплекс прикрепляется к актиновым филаментам и придает актомиозину скелетных мышц позвоночных чувствительность к ионам Са2+ (рис. 20.6). Установлено, что тропонин (его субъединицы Тн-Т и Тн-I) способен фосфорилироваться при участии цАМФ-зависимых протеинкиназ. Вопрос о том, имеет ли отношение фосфорилирование тропонина in vitro к регуляции мышечного сокращения, остается пока открытым. Белки стромы в поперечно-полосатой мускулатуре представлены в основном коллагеном и эластином. Известно, что строма скелетных мышц, остающаяся после исчерпывающей экстракции мышечной кашицы солевыми растворами с высокой ионной силой, состоит в значительной мере из соединительнотканных элементов стенок сосудов и нервов, а также сарколеммы и некоторых других структур.
Рис. 20.6. Структура тонкого филамента. 1 - актин; 2 - тропомиозин; 3 - тропонин С; 4 - тропонин I; 5 - тропонин Т.
137.Особенности химического состава и метаболизма нервной ткани. 2. ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ В НЕРВНОЙ ТКАНИ. 3. ХИМИЧЕСКАЯ ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ. ХИМИЧЕСКИЙ СОСТАВ НЕРВНОЙ ТКАНИ. Функции нервной системы: 1. координация и регуляция обмена веществ, 2. обеспечение связи организма с внешней средой. Химический состав нервной ткани сложен и неоднороден, как в целом и сама нервная ткань. Отличия в химическом составе носят в основном количественный характер. В сером веществе 77 -81% воды, в белом-70%. Содержание белков в нервной ткани меньше, чем в некоторых других тканях (печень, мышцы). Их больше в сером веществе и меньше в периферических нервах. В нервной ткани содержится больше сложных белков: ЛИПОПРОТЕИНЫ (миелиновые оболочки), ФОСФОПРОТЕИНЫ, НУКЛЕОПРОТЕИНЫ (ДНП, РНП), ГЛИКОПРОТЕИНЫ (нейрокератин), En. Наибольший интерес представляют нейроспецифические белки: 1. белок S-100 (растворим в 100% растворе (NH4)2SO4). Он повышается в ткани мозга в эксперименте при обучении и эмоциональном воздействии. Считают, что этот белок отвечает за формирование зависимостей (алкогольной, наркотической). ПРОПРОТЕИН - антитело к белку S-100, понижает его содержание в ткани мозга. 2. Белок 14-3-2 играет важную роль в формировании памяти. 3. НЕЙРОПЕПТИДЫ - играют роль нейромедиаторов и гормонов. Пептиды памяти, боли, сна. Не белковые азотистые соединения те же самые, что и в других тканях, но отличаются по количественному составу. В нервной ткани много свободных аминокислот, г.о. дикарбоновых (ГЛУ, ГЛН, ACП, АСН), ГАМК, ароматические аминокислоты, ЦАМФ и ЦГМФ. Углеводов незначительное количество:
1. резервные углеводы - гликоген (0,1 %) 2. глюкоза (1 -4мкмоль/л) 3. гексозофосфаты 4. триозофосфаты 5. ПВК 6. молочная кислота. ЛИПИДОВ в сером веществе 25%, в белом веществе 50%. 1. ФОСФОЛИПИДЫ (до 50%) - ПЛАЗМОГЕН АЦЕТАЛЬФОСФАТИДЫ. 2. ГЛИКОЛИПИДЫ - ЦЕРЕБРОЗИДЫ, ГАНГЛИОЗИДЫ. 3. ВЖК - в основном непредельные, содержащие по 4 - 5 двойных связей. 4. Холестерин (25%) в свободном виде. Мозг даже называют депо холестерина. 5. Нейтральные жиры - в незначительном количестве в головном, но в большом количестве в периферических нервах. Минеральные вещества представлены катионами калия, натрия, кальция, магния, железа, меди, цинка, в качестве анионов выступают анионы белков и фосфаты. ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ В НЕРВНОЙ ТКАНИ. Особенности энергетического обмена. 1. В ткани головного мозга очень высока интенсивность дыхания. Мозг потребляет кислород больше, чем постоянно функционирующий миокард, на 20% больше покоящиеся мышцы. 20 - 25% всего кислорода приходится на долю головного мозга, у детей грудного возраста -50%. За 10 секунд мозг поглощает весь кислород, находящийся в ткани. Из этого следует особая важность адекватного кровоснабжения нервной ткани. Прекращение кровоснабжения ГМ 6 -8 секунд приводит к потере сознания. 2. Основным субстратом окисления является глюкоза. ДЫХ. КОЭФ.=V(СО2)/V(О2)=1,0 Мозг это единственный орган, использующий в качестве источника энергии исключительно глюкозу. 3. До 70% АТФ используется для поддержания ионных градиентов (удаление натрия из клетки). ОСОБЕННОСТИ УГЛЕВОДНОГО ОБМЕНА.
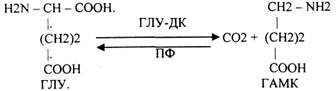
Исходный субстрат окисления - глюкоза. 85% окисляется аэробно. Функция головного мозга зависит от концентрации глюкозы в крови. ГИПОГЛИКЕМИЯ ведёт к судорогам, смерти. 0,1% гликогена обновляется в ткани мозга за 4 часа. Распад идет двумя путями: ФОСФОРОЛИТИЧЕСКИМ и ГИДРОЛИТИЧЕСКИМ. При нарушении обмена углеводов нарушается функция головного мозга. Эффект снотворных веществ связан с торможением окисления углеводов. Недостаток витамина BI ведёт к нарушению углеводного обмена и полиневритам. ОСОБЕННОСТИ ОБМЕНА БЕЛКОВ И АМИНОКИСЛОТ. При возбуждении нервной ткани нарушается распад белков и аминокислот и повышается содержание аммиака в клетках. Во время торможения и сна количество аммиака снижается. Аммиак токсичен для ткани головного мозга и обезвреживается ГЛУ с образованием амидов (ГЛН)
ГПН проникает через мембраны и выходит из клетки. РОЛЬ ГЛУТАМИНОВОЙ КИСЛОТЫ В ОБМЕНЕ ВЕЩЕСТВ. 1. ГЛУ связывает аммиак 2. ТРАНСАМИНИРОВАНИЕ, следовательно, образование заменимых аминокислот 3. ДЕКАРБОКСИЛИРОВАНИЕ
4 ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
5. Является нейромедиатором возбуждающего действия 6. Стабилизирует содержание калия в клетке. До 10% глюкозы превращается в нервной ткани в ГЛУ. ГЛУ применяется в качестве лечебного средства при ЧМТ. ОСОБЕННОСТИ ЛИПИДНОГО ОБМЕНА. Нейтральные жиры не используются как энергетический материал в нервной ткани. ФОСФО и ГЛИКОЛИПИДЫ играют в основном структурную роль, как компоненты мембран. ХИМИЧЕСКАЯ ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ. Химическими передатчиками являются нейромедиаторы: 1. АЦЕТИЛХОЛИН 2. БИОГЕННЫЕ АМИНЫ 3. АМИНОКИСЛОТЫ (ГЛУ, ГЛИ) 4. НЕЙРОПЕПТИДЫ. В передаче нервного возбуждения участвуют: 1. Натриевый насос (АТФ -аза) 2. Натриевые каналы 3. Калиевые каналы. Этапы передачи нервного возбуждения. 1. Нервный импульс 2. Освобождение медиатора 3. Диффузия медиатора через синаптическую щель 4. Присоединение медиатора к рецептору 5. Изменение конфигурации рецептора 6. Открытие калий -натриевых каналов 7. Инактивация медиатора 8. Удаление натрия из клетки с помощью АТФ-азы. В покое натрий удаляется из клетки, и мембрана становится проницаемой для калия. Калий выходит из клетки и внутри клетки наблюдается положительный потенциал - ПП (70мВ) При возбуждении натрий -калиевые каналы открываются, натрий заходит в клетку, калий выходит из клетки. Но т.к. концентрация натрия снаружи больше, его в клетку заходит больше. Это приводит к понижению отрицательного заряда внутри клетки и мембрана деполяризуется. Это и есть начало передачи импульса по мембране. ОБРАЗОВАНИЕ НЕЙРОМЕДИАТОРОВ. В ХОЛИНЕРГИЧЕСКИХ синапсах основной медиатор - АЦЕТИЛХОЛИН. Образуется из холина.
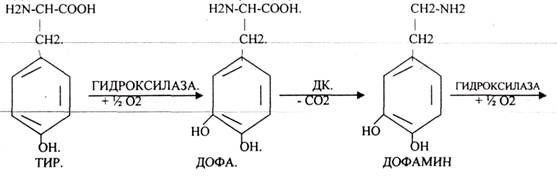
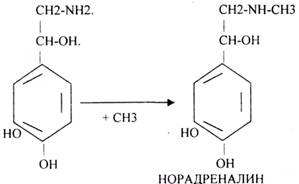
Ацетилхолин разрушается с помощью ХОЛИНЭСТЕРАЗЫ. В АДРЕНЕРГИЧЕСКИХ синапсах медиаторы образуются из ТИР.
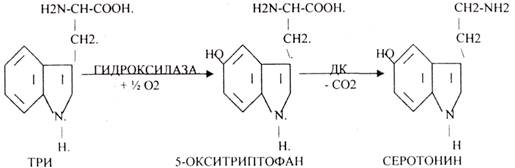
В СЕРОТОНИНЕРГИЧЕСКИХ синапсах медиатор образуется изТРИ.
Они разрушаются МОНОАМИООКСИДАЗОЙ (МАО).
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 53; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.196.217 (0.077 с.) |