Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Г люкозо - лактатный и глюкозо - аланиновый циклы
Глюкозо-лактатный цикл (цикл Кори) –это связь глюконеогенеза в печени и об-
разования лактата эритроцитах или мыш-цах из глюкозы. В эритроцитах молочная
кислота образуется
непрерывно, так как для них анаэробный
гликолиз является
единственным спосо-бом образования энер-
гии. В скелетных мыш-цах накопление лакта-та является следстви-ем гликолиза при очень интенсивной, максимальной мощно-
сти, работе, и чем более такая работа интенсивна, тем менее продолжительна По-
сле нагрузки (во время восстановления) лактат удаляется из мышцы довольно бы-
стро – всего за 0,5-1,5 часа.
Дополнение
Следует отметить, что если продолжи-
тельность нагрузки мала (до 10 секунд), то количество АТФ пополняется пре-имущественно в креатинфосфоки-назной реакции. В
таком режиме к при-меру работают мышцы у ттанги- стов, прыгунов как в длину, так и в высо- ту, метателей мо-лота, копья и т.п.. Если нагрузка не бо-лее 90 секунд – АТФ синтезируется в основном в реакциях анаэробного глико- лиза. В спорте это бегуны-спринтеры на 100-500 м, спортсмены силовых ви-дов (борцы, тяжелоатлеты, бодибилдеры). Если напряжение мышцы длится более двух минут – развивается аэробное окисление глюкозы в реакциях ЦТК и дыхательной цепи.
Но, хотя мы и говорим об аэробном окислении глюкозы, необходимо знать и помнить, что лактат образуется в мышце всегда: и при анаэробной, и при аэробной работе, однако в разных количествах.
Образовавшийся лактат может утилизоваться только одним способом – превра-
титься в пировиноградную кислоту. Но, как уже указывалось, пируват токсичен для
клеток и должен быть как можно быстрее утилизован. Сама мышца ни при работе, ни во время отдыха не занимается превращением лактата в пируват из-за наличия
специфического изофермента ЛДГ-5.
Если молочная кислота поступила в миокардиоциты, она быстро превраща-ется в пируват, далее в ацетил- S -КоА и вовлекается в полное окисление до
СО B 2 B и Н B 2 B О.
Большая часть лактата захватывается гепатоцитами, окисляется в пировино-
градную кислоту и вступает на путь глюконеогенеза.
Целью глюкозо-аланинового цикла также является уборка пирувата, но, кроме
этого решается еще одна немаловажная задача – уборка лишнего азота из мышцы.
При мышечной работе и в покое в миоците распадаются белки и образуемые амино-кислоты рансаминируются с α-кетоглутаратом. Полученный глутамат взаимодейст-
вует с пируватом. Образующийся аланин является транспортной формой и пирувата
и азота из мышцы в печень. В гепатоците идет обратная реакция трансаминирова-ния, аминогруппа передается на синтез мочевины, пируват используется для синте-за глюкозы
Глюкоза, образованная в печени из лактата или аланина, возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время го-
лодания, когда мышечные белки распадаются и многие аминокислоты используются
в качестве источника энергии, а их азот необходимо доставить в печень.
Р ЕГУЛЯЦИЯ ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА
МЕТАБОЛИЗМ ЭТАНОЛА
С ПИРТОВОЕ БРОЖЕНИЕ
Образование этилового спирта из глюкозы происходит в дрожжах и некоторых видах плесневых грибков. Суммарное уравнение реакции: C6H5О10 → 2 CО2 + 2 С2Н5ОН
До стадии образования пирувата реакции спиртового броже-
ния совпадают с реакциями гликолиза, отличия заключаются
только в дальнейшем превращении пировиноградной кислоты. Цель этих превращений – удалить пируват из клетки и окислить
НАДН, который образовался в 6-й реакции.
О БЕЗВРЕЖИВАНИЕ ЭТАНОЛА
Метаболизм поступающего этанола в организме происходит
в печени двумя путями. Первый путь заключается в окислении
спирта до уксусной кислоты, которая в виде ацетил-S-КоА посту-
пает в ЦТК. Через этот путь проходит от 70% до 90% всего эта-нола. Оставшаяся часть окисляется в микросомах алкогольокси-дазой. При регулярном поступлении этанола доля микросомаль-ного окисления возрастает, количество молекул алкогольоксида-зы увеличивается.
Так как при обезвреживании этанола образуется большое количество НАДН, в клетках печени активируется реакция превращения пирувата в лактат. Это приводит к гипогликемии, так как пировиноградная кислота является субстратом глюконео-генеза. Свободное проникновение молочной кислоты в кровь обуславливает лакта-
цидемию.
Если запасы гликогена в печени изначально невелики (голодание, недоедание, астеническое телосложение) или израсходованы (после физической работы), то при приеме алкоголя натощак гипогликемия наступает быстрее и может быть причиной потери сознания. К этому стоит добавить сильный диурети- ческий эффект этанола, ведущий к быстрому обезвоживанию организма и снижению кровоснабжения головного мозга со всеми вытекающими последст-виями.
Этанол является энергетически ценным соединением: при метаболизме 125 г этанола количество образующегося НАДН такое же, как при окислении 500 г глюко- зы. При полноценном питании и частом потреблении этилового спирта, например, в виде пива, "этанольный" ацетил-S-КоА не столько сгорает в ЦТК, сколько использу- ется для синтеза холестерина и нейтральных жиров, то есть происходит переход
энергии этанола в запасную форму, что приводит к пивному ожирению и повышает риск атеросклероза.
ПЕНТОЗОФОСФАТНЫЙ ПУНТЬ
Пентозофосфатный путь окисления глюкозы не связан с образованием энергии.
Значение ПФП:
1. Образование НАДФН
• для синтеза жирных кислот,
• холестерина и других стероидов,
• для синтеза глутаминовой кислоты из α-кетоглутаровой кислоты (реак-
ция восстановительного аминирования).
• для систем защиты клетки от свободно-радикального окисления (анти-оксидантная защита).
2. Образование рибозо-5-фосфата, необходимого для синтеза нуклеиновых ки-
слот.
Наиболее активно реакции ПФП идут в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, менее интенсив- но в скелетных мышцах.
Пентозофосфатный путь включает два этапа – окислительный и неокислитель-ный. На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превра-
щается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух мо-лекул НАДФ до НАДФН. Второй этап – этап структурных перестроек, благодаря которым пентозы воз-
вращаются в фонд гексоз. В этих реакциях рибулозо-5-фосфат может изомеризо-ваться до рибозо-5-фосфата и ксилулозо-5-фосфата. Далее под влиянием фермен- тов транскетолазы и трансальдолазы происходят структурные перестройки с обра-
зованием иных моносахаридов. При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегидфосфат. Из глицеральдегид- 3-фосфата при необходиости могут образоваться гексозы.
Связь пентозофосфатного пути и гликолиза
Судьба полученных фруктозо-6-фосфата и глицеральдегидфосфата различна
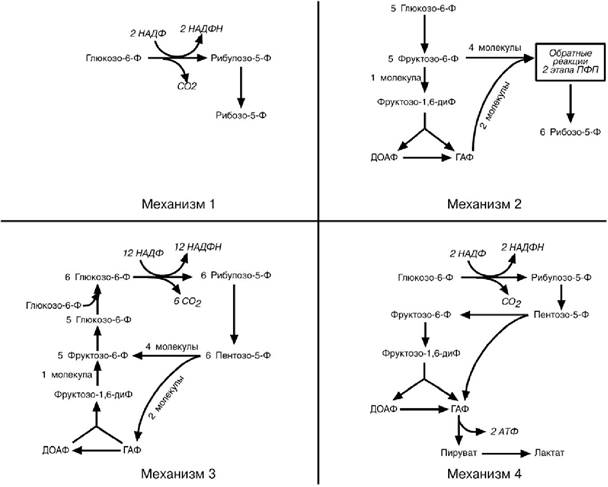
в зависимости от ситуации и потребностей клетки. Поэтому метаболизм глюкозо-6-фосфата может идти по 4 различным механизмам.
Механизм 1. Потребность в НАДФН и рибозо-5-фосфате сбалансирована
(например, при синтезе дезоксирибонуклеотидов). При таких условиях реак-
ции идут обычным порядком – образуется две молекулы NADPH и одна моле-
кула рибозо-5-фосфата из одной молекулы глюкозо-6-фосфата по окисли-тельной ветви пентозофосфатного пути.
Механизм 2. Потребность в рибозо-5-фосфате значительно превышает потребность в НАДФН (например,синтез РНК)Большая часть глюкозо-6-фосфата превращается во фруктозо-6-фосфат и глицеральдегид-3-фосфат
по гликолитическому пути. Затем две молекулы фруктозо-6-фосфата и одна молекула глицеральдегид-3-фосфата под действием трансальдолазы и транскетолазы рекомбинируют в три молекулы рибозо-5-фосфата путем обращения реакции 2 этапа пентозофосфатного пути.
Механизм 3. Потребность в НАДФН значительно превышает потреб-
ность в рибозо-5-фосфате (например,биосинтез холестерола,жирных ки-слот). В этой ситуации по окислительным реакциям пентозофосфатного пути образуются НАДФН и рибулозо-5-фосфат. Далее, под действием транс-кетолазы и трансальдолазы, рибулозо-5-фосфат превращается в пентозо-5-фосфаты, во фруктозо-6-фосфат и глицеральдегид-3-фосфат. В заключение происходит ресинтез глюкозо-6-фосфата из фруктозо-6-фосфата и глице-ральдегидфосфата по пути глюконеогенеза. Подключение новых молекул глюкозо-6-фосфата позволяет поддерживать стехиометрию процесса.
Механизм 4. Потребность в НАДФН значительно превышает потреб-ность в рибозо-5-фосфате и необходима энергия (например.антиокси-
дантная защита в эритроците). Глюкозо-6- фосфат превращается в рибозо-5-фосфат и далее во фруктозо-6- фосфат и глицеральдегид-3-фосфат, ко-торые (в отличие от механизма 3) вступают на гликолитический путь обме-на, а не подвергаются обратному превращению в глюкозо-6-фосфат. Образо-ванный пируват вступает в ЦТК. В результате происходит одновременное генерирование НАДФН и АТФ.
|
|||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 134; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.147.215 (0.03 с.) |