Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности свойств сердечной мышцы и ее энергетического обеспечения
Рабочему миокарду сердца присущи те же свойства, что и скелетной мышце, но есть существенные особенности их. Особенности возбудимости и возбуждения сердечной мышцы Ионный механизм возникновения ПД кардиомиоцитов представлен на рис. 11.3. Фаза деполяризации и восходящая часть фазы инверсии осуществляются в основном за счет входа Na+ в клетку, как у скелетных миоцитов. В этот период увеличена проницаемость мембраны для Na+ в связи с открытием быстрых Na-каналов, когда деполяризация достигает КП (примерно —40 мВ), Na+ лавиной входит в клетку. В фазу деполяризации Na+ входит в клетку согласно концентрационному и электрическому градиентам (клетка в эту фазу внутри еще имеет отрицательный заряд). В восходящую часть фазы инверсии Na+ входит в клетку только согласно концен-
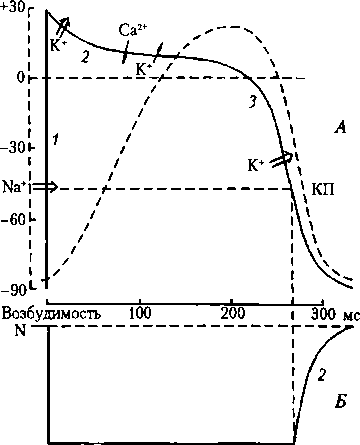
Рис. 11.3. Механическая и электрическая активность сердца:
А — потенциал действия (сплошная линия), сокращение миокарда (пунктир): 1 — фаза деполяризации, 2 — фаза инверсии, 3 — фаза реполяризации (-> — медленный ток ионов, => — быстрый ток ионов); Б — фазовые изменения возбудимости клетки рабочего миокарда в процессе ее возбуждения: N — нормальная возбудимость, 1 — фаза абсолютной рефрактерное™, 2 — фаза относительной рефрактерности трационному градиенту, но вопреки электрическому — в этот период клетка перезаряжается: внутри клетки формируется положительный заряд, снаружи — отрицательный. Нарастание ПД прекращается вследствие инактивации быстрых Na-каналов. Если в эксперименте уменьшить концентрацию Na+ вне клетки со 140 до 20 мМ, клетка становится невозбудимой. Нисходящая часть фазы инверсии (плато) начинается в результате активации К-каналов и выхода К+ из клетки. Вначале спад ПД происходит быстро, вследствие быстрого выхода К+ из клетки, так как в это время электрический градиент К+ максимальный (клетка внутри заряжена положительно), совпадающий с концентрационным градиентом К+. Далее проводимость К+ уменьшается, поэтому уменьшается выход К+ из клетки, активируются Са-каналы и Са2+ поступает в клетку, в результате чего спад ПД замедляется. Затем медленный суммарный вход Са2+ в клетку становится примерно равным медленному выходу К+ из клетки, что обеспечивает возникновение плато ПД в нисходящей части фазы инверсии. Медленный спад плато свидетельствует о том, что выход К+ из клетки несколько превышает вход Са2+ в клетку вследствие начинающейся инактивации медленных Са-каналов (L-типа; англ, long— долгий).
Фаза реполяризации начинается вследствие инактивации медленных Са-каналов и активации быстрых К- каналов, в результате чего К+ начинает лавиной выходить из клетки, что обеспечивает развитие фазы реполяризации кардиомиоцитов. Мембранный потенциал возвращается к исходной величине -85-90 мВ. В фазу инверсии (нисходящая часть) К+ выходит из клетки согласно концентрационному и электрическому градиентам, в фазу реполяризации — согласно концентрационному, но вопреки электрическому — в эту фазу клетка снаружи уже снова заряжена положительно, а изнутри — отрицательно. Са2+ из кардиомиоцита после ПД выносится главным образом Ca/Na- обменником, частично — Са-АТФазой. Параметры сердечных потенциалов. Формируется ПП клеток рабочего миокарда в основном градиентом К+, его величина составляет —85— 90 мВ. Амплитуда ПД 120 мВ; длительность ПД кардиомиоцитов желудочков (300—350 мс) почти соответствует длительности сокращения мышцы сердца. Продолжительность ПД кардиомиоцитов предсердия — до 120 мс, почти столько же длится и систола предсердий. Длительность абсолютной рефрактерной фазы кардиомиоцитов желудочков примерно в 200—300раз больше (см. рис. 11.3, Б) таковой миоцитов скелетной мышцы. У скелетной мышцы пик ПД длится 1—3 мс. Примерно столько же длится и абсолютная рефрактерная фаза, которая соответствует длительности пика ПД, поскольку во время пика имеет место инактивация быстрых Na-каналов. Так же и кардиомиоциты — пока длится ПД, они невозбудимы, что связано в основном с инактивацией быстрых Na-каналов. Период абсолютной реф- рактерности для миокарда желудочков составляет 270 мс, он соответствует фазе инверсии (плато ПД) и начальной части фазы реполяризации до достижения КП (около —40 мВ); период относительной рефрактерности (около 30 мс) соответствует остальной части фазы реполяризации (см. рис. 11.3, Б). Длительная рефрактерная фаза предотвращает круговое распространение возбуждения по миокарду и исключает тетаническое сокращение миокарда.
Характеристика проводимости сердечной мышцы Возбуждение в сердечной мышце распространяется диффузно, что объясняется особенностями ее строения. Миокард, хотя он и состоит из отдельных клеток, как и скелетная мышца, функционирует как единое целое — функциональный синцитий (возбуждение распространяется от одной клетки к другой во всех направлениях за счет электрического поля возбужденной клетки к невозбужденной). Клетки сердечной и гладкой мышц, глии и амакринные клетки сетчатки глаза соединены между собой щелевыми контактами (щель — около 2 нм). Кардиомиоциты соединяются друг с другом своими концами с помощью вставочных дисков, являющихся продолжением сарколемм клеток. В области вставочных дисков имеются высокопроводимые щелевые контакты (в сердце их называют нексусы), с помощью которых возбуждение передается от одной клетки к другой. Кардиомиоциты имеют цилиндрическую форму (их длина 100-150 мкм, диаметр 20 мкм), могут ветвиться и образовывать пространственную сеть. Боковые поверхности кардиомиоцитов также соединены между собой нексусами. Они построены из белковых субъединиц — коннексинов, образующих комплекс, называемый коннексоном. Коннексон одной клетки контактирует с коннексоном соседней клетки, образуя канал между двумя клетками. Такие соединения в возбудимых клетках называют также электрическими синапсами. Считается, что возбуждение от одной клетки к другой в этом случае проводится благодаря движению ионов: катионов из возбужденной клетки к невозбужденной (заряд последней внутри отрицательный), а анионов — в противоположном направлении. Время проведения возбуждения через один электрический синапс (задержка) очень короткое — 10-5с. По мнению, например, Rainer Klinke (2004), описанный механизм проведения возбуждения через щелевые контакты похож на таковой в немиелинизирован- ном нервном волокне. Следует, однако, заметить, что время задержки не раскрывает механизм передачи сигнала. Причем если данное время очень мало, то это доказывает не ионный, а электрический механизм передачи сигнала — возникший ПД в одной клетке возбуждает соседнюю клетку за счет своего электрического поля. В этом случае время затрачивается не на движение ионов от клетки к клетке, а только на возникновение ПД, т.е. перпендикулярно относительно клеточной мембраны — в клетку и из клетки. Однако, согласно нашим расчетам, эта задержка (10-5 с) очень велика. Если бы возбуждение проходило от одной клетки к другой (щель между ними примерно 2 нм и толщина двух мембран по 4 нм) за 10-5 с, то на прохождение 1 м пути потребовалось бы 1000 с, что в 1 тыс. раз медленнее реальной скорости (1 м/с). Таким образом, клетки организма взаимодействуют между собой с помощью веществ и электрических полей. Так, возбуждение распространяется в пределах одной клетки (по нервным волокнам и миоцитам) с помощью электрического поля ПД. К другой клетке сигнал передается посредством веществ (в химических синапсах) или электрического поля (в электрических синапсах). ЛП (ВПСП, РП и ГП) обеспечивают возбуждение нейронов и миоцитов (исчерченных и гладких мышц) также посредством действия на них своих электрических полей. При этом ТПСП и ТПД модулируют взаимодействие клеток организма. При исследовании электрической активности головного мозга, сердца, желудка и мышц с помощью накожных электродов также регистрируется суммарное электрическое поле ПД и ЛП клеток этих органов. Скорость проведения возбуждения по миокарду (около 1 м/с), примерно в 3,5 раза меньше, чем у скелетной мышцы, и в 100 раз меньше, чем в нервном волокне.
|
||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 175; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.171.121 (0.01 с.) |