Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нейронная организация новой коры. Кортикализация функций
Новая кора (неокортекс) — это слой серого вещества, общая площадь которого достигает 2 тыс. см2 за счет складок; неокортекс покрывает большие полушария и составляет около 70 % всей площади коры. В направлении с поверхности вглубь неокортекс имеет 6 горизонтальных слоев (см. рис. 6.1),»архиокортекс — 3, па- леокортекс — 4—5. Функциональные слои новой коры большого мозга I. Молекулярный слой имеет мало клеток, но содержит большое количество ветвящихся, восходящих дендритов пирамидных клеток, на которых образуют синапсы волокна, приходящие от ассоциативных и неспецифических ядер таламуса и регулирующие уровень возбудимости коры. II. Наружный зернистый слой содержит в основном звездчатые клетки и, частично, малые пирамидные клетки. Волокна его клеток располагаются преимущественно вдоль поверхности коры, образуя кортико-кортикальные связи. III. Пирамидный слой сформирован в основном из пирамидных клеток средней величины, аксоны которых образуют кортико-кортикальные ассоциативные связи, как и зернистые клетки II слоя. IV. Внутренний зернистый слой образован звездчатыми клетками, на которых имеются синапсы от волокон нейронов специфических ядер таламуса и метаталамуса, несущих информацию от рецепторов сенсорных систем. V. Ганглионарный слой представлен средними и крупными пирамидными клетками. Причем гигантские пирамидные клетки Беца расположены в двигательной коре, их аксоны образуют пирамидные пути — кортикобульбарный и кортикоспинальный двигательные пути (пирамидные пути). VI. Слой полиморфных клеток, аксоны которых образуют кортикоталамические пути. В слоях I и IV новой коры происходит восприятие и обработка поступающих сигналов. Нейроны II и III слоев осуществляют кортико-кортикальные ассоциативные связи. Нейроны V и VI слоев формируют нисходящие пути. Функциональные нейронные колонки новой коры большого мозга. В коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5-1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов (нейронные колонки). Об этом, в частности, свидетельствуют электрофизиологические исследования В. Маунткасла (1957) с погружением микроэлектродов перпендикулярно к поверхности соматосенсорной коры. При этом все встречаемые на пути нейроны отвечают на раздражитель только одного вида (например, свет). При погружении электрода под углом на его пути попадались нейроны разной сенсорности. Колонки обнаружены в моторной коре и различных зонах сенсорной коры. Нейроны колонки могут осуществлять саморегуляцию по типу возвратного торможения. Соседние нейронные колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения.
Кортикализация функций. Под корти- кализацией функций понимают возрастание в филогенезе роли коры большого мозга в регуляции функций организма и подчинение себе нижележащих отделов ЦНС, в обеспечении психической деятельности организма. Например, регуляция локомоторных двигательных функций (прыжки, ходьба, бег) и выпрямительных рефлексов у низших позвоночных полностью обеспечивается стволом мозга, и удаление больших полушарий практически их не изменяет. У кошек после перерезки ствола между средним и промежуточным мозгом локомоция сохраняется лишь частично. Выключение коры большого мозга в эксперименте у обезьян и в патологических случаях у человека ведет к потере не только произвольных движений, локомоции, но и выпрямительных рефлексов. 6.22. Основные функциональные зоны новой коры Общая характеристика коры. В новой коре имеются поли- и монофункциональные поля. Свойство полифункциональности позволяет данной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию (Андрианов О. С., 1976). Степень полифункциональности различных корковых структур неодинакова; например, поля ассоциативной коры по- лифункциональны. Первичные сенсорные зоны моносенсорны. Во вторичных сенсорных зонах локализуются преимущественно бисенсорные нейроны. В основе мультифункциональности лежит многоканальность поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений, особенно на таламическом и корковом уровнях, модулирующее влияние различных структур (например, неспецифического таламуса, базальных ядер) на корковые функции, взаимодействие корково-подкорковых и межкорковых путей проведения возбуждения.
Детальное деление коры на различные поля проведено на основе цитоар- хитектонических признаков (формы и расположения нейронов) К. Бродманом (1909), выделивший 52 поля, многие из которых характеризуются функциональными и нейрохимическими особенностями. Более крупным вариантом функционального разделения коры головного мозга является выделение в ней сенсорной, ассоциативной и двигательной зон. Сенсорные зоны коры (проекционная кора, корковые отделы анализаторов сенсорных систем) Характеристика сенсорной к о р ы. В эти зоны поступает информация от сенсорных рецепторов. Они расположены в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса. Участки сенсорной коры,
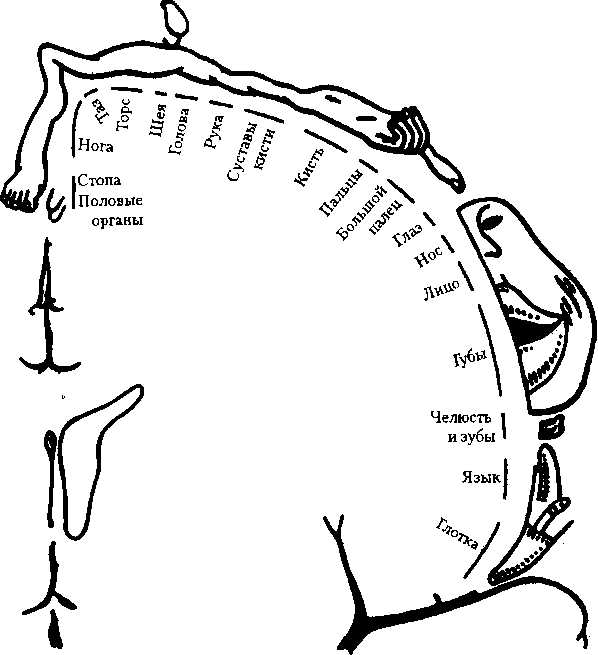
Соматосенсорная область (S,) — это постцентральная извилина теменной доли и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1-3). При этом поле 3 представляет собой первичное сенсорное поле, а поля 1-2 — вторичные сенсорные поля. В области Sj имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и проприоцептивной чувствительности (рис. 6.21). Проекции наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеют относительно большие зоны по сравнению с другими частями тела. Предполагается, что в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности. Имеется (также меньших размеров) вторичная соматосенсорная область (Sn), расположенная в стенке боковой борозды, на границе пересечения последней с центральной бороздой. Лобная и теменная доли, как известно, разделены центральной бороздой. Зона S,,, по-видимому, участвует в сенсорной и моторной координации двух сторон тела; локализация поверхности тела в ней менее четкая, чем в области Sr Височная область. Здесь в верхней височной извилине расположена слуховая сенсорная кора. Первичной сенсорной зоной является поле 41, в глубине боковой борозды с четкой топической проекцией различных участков кортиева органа. Вторичная сенсорная зона расположена в верхней височной извилине (поле 42), где происходит более сложная обработка звуковой (частично и речевой) информации.
Затылочная область (поле 17). Здесь локализуется первичная сенсорная зона зрительной системы с топическим представительством рецепторов сетчатки, особенно желтого пятна (основная часть поля 17), рядом располагается вторичная сенсорная зона (поля 18 и 19). Обработанная в первичной и вторичной сенсорной коре информация передается для дальнейшей переработки в ассоциативную кору — третичную сенсорную зону. Ассоциативные области коры Характеристика ассоциативной коры. Ассоциативная кора включает участки новой коры большого мозга, которые расположены рядом с сенсорными и двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у человека, составляя у него 70-75 % не- окортекса. Границы этих областей обозначены недостаточно четко в связи с плавным переходом свойства вторичных сенсорных полей (где локализуются преимущественно бисенсорные нейроны) к полисенсорным полям. Полисенсорность нейронов ассоциативной коры создается как ее кортикокортикальными связями с разными проекционными зонами, так и главным ее афферентным входом от ассоциативных ядер таламуса, в которых уже произошла обработка информации от различных рецептивных полей. В результате обработки информации в ассоциативных областях, взаимодействующих с первичными и вторичными сенсорными зонами, формируется целостное представление об окружающей действительности. Совокупность ассоциативных ядер таламуса и ассоциативных зон коры выделяют как ассоциативные системы мозга: таламолобную, таламотеменную и таламовисочную. Следует, однако, заметить, что ассоциативные зоны (как третичные сенсорные области) без взаимодействия с первичными и вторичными сенсорными зонами обеспечить восприятие не могут. Поэтому понятие ассоциативная система мозга должно включать ассоциативные области коры, переключательные и ассоциативные ядра таламуса, а также первичные и вторичные зоны коры, а восприятие — формирование образа объекта или явления, действующих на органы чувств (это одна из форм психической деятельности). При этом происходит узнавание ранее встречаемых объектов, явлений или формирование нового образа — впервые встретившихся объектов или явлений. Узнавание же возможно только с помощью сличения поступающей в данный момент информации с закодированной с помощью механизмов памяти информацией (со следами памяти). Таким образом, можно выделить три основные сенсорно-ассоциативные системы головного мозга: 1) таламотеменную; 2) таламовисочную; 3) таламо- затылочную. Каждая из них включает соответствующие ядра таламуса, первичную и вторичную сенсорные зоны, а также ассоциативную кору — третичную сенсорную зону.
Таламотеменная система, кроме ядер таламуса, первичной и вторичной сенсорных зон, включает ассоциативную зону теменной коры (поля 5, 7, 39 и 40). Она имеет эфферентные выходы на моторную кору и ядра экс- трапирамидной системы. Основными ее функциями являются гнозис и прак- сис. Гнозис — узнавание формы, величины, значения предметов, оценка их пространственных отношений, понимание речи, познание процессов, закономерностей и др. Например, в теменной коре выделяют центр стереогнозиса, обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»). Праксис — целенаправленное действие. Ведущую роль в праксисе играет надкраевая извилина (поля 39 и 40) — хранение и реализация программ двигательных автоматизированных актов. Таким образом, теменная кора участвует в осознании (восприятии) соматосенсорной информации, влияет на произвольное движение посредством лобной доли, через нее реализуется зрительный контроль произвольных движений, она участвует в восприятии пространства. Тал амо в и с очная система образована по тому же принципу. Височная доля обеспечивает восприятие звуковых сигналов, переработку вестибулярной информации, участвует в переработке зрительной информации (например, нижняя височная извилина участвует в распознавании лиц), включает речевой центр Вернике. Медиальная часть височной доли, являясь элементом лимбической системы, участвует в регуляции вегетативных функций и в формировании эмоций и поведения организма. Гиппокампальная формация (медиальная часть височной доли), включающая гиппокамп (аммонов рог), зубчатую извилину, суби- кулум (основание гиппокампа), играет важную роль в формировании памяти и в научении (см. п. 18.2, 18.3). Таламозатылочная система обеспечивает зрительное вое- приятие, участвует также в управлении движениями глаз, в аккомодации, в осуществлении зрачковых реакций. Лобная кора не содержит первичных и вторичных сенсорных зон. Главной функцией лобной ассоциативной коры (поля 9-14) является инициация движения и его план, особенно в новой для человека обстановке. В случае поражения лобной ассоциативной коры или в результате лоботомии, при которой по медицинским показаниям пересекаются связи между лобной долей и таламусом, наблюдается развитие «эмоциональной тупости», отсутствие мотивации, твердых намерений и планов, основанных на прогнозировании. Такие люди становятся грубыми, у них появляется тенденция к повторению каких-либо привычных, не соответствующих обстановке двигательных актов, нарушается внимание, принятие решений, социальное поведение.
Таким образом, лобная доля (премоторная дополнительная двигательная область, глазодвигательное поле, двигательный центр речи — центр Брока) осуществляет планирование и выполнение движений, участвует в формировании эмоций, личностных качеств человека. Двигательные зоны коры — это первичная двигательная (моторная) кора прецентральной извилины (поле 4); вторичная двигательная кора (поле 6), которая включает прилегающую к ней рострально премоторную область и дополнительную двигательную область на медиальной стороне полушария, а также вторичную соматосенсорную кору на крыше латеральной щели (область Sn). Пирамидные нейроны V слоя этих областей иннервируют а- и у-мотоней- роны двигательных центров как непосредственно (моносинаптически), так и через интернейроны двигательных центров (полисинаптически), составляют двигательные колонки. Пирамидные нейроны двигательной колонки получают импульсы от проприо- и тактильных рецепторов, формируют двигательные команды и посылают их через нейроны верхних слоев к стволовым и спинальным центрам; последние управляют мышцами одного сустава или одной мышцы. Однако мышцы туловища могут иннервироваться от нескольких колонок. Соседние колонки в функциональном плане перекрываются. В двигательные области информация поступает по восходящим афферентам через таламус и интегрированная информация от корковых нейронов различных сенсорных систем (например, проприоцептивная, соматосенсорная) и от ассоциативной коры и лимбической системы. Таким образом, двигательная кора представляет собой общий конечный путь в организации движений. Основной эфферентный выход двигательной коры на стволовые и спинальные моторные центры формируют пирамидные клетки Беца и менее крупные пирамидные клетки коры прецентральной извилины (60 % волокон), премоторной коры (20 % волокон) и постцентральной извилины (20 % волокон). Двигательная кора — организатор движений. Первичная моторная кора. Пирамидные клетки первичной моторной коры иннервируют мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (рис. 6.22). Проекции мышц нижних конечностей и туловища расположены в верхних участках прецентральной извилины и занимают сравнительно небольшую площадь, а проекции мышц верхних конечностей, лица и языка, обеспечивающих наибо Вторичная двигательная кора имеет главенствующее значение по отношению к первичной двигательной коре и обеспечивает осуществление более сложных координированных движений, играет важную роль в планировании движения. Поражение вторичной двигательной зоны ведет к резкому ограничению движений у человека.
Как отмечалось выше, двигательная кора осуществляет свои функции через пирамидную и экстрапирамидную системы, связанные с одними и теми же мотонейронами двигательных центров черепных нервов и спинного мозга. Пирамидная система — это совокупность пирамидных клеток V слоя коры большого мозга (клетки Беца), «-мотонейронов черепных нервов (III—VII и IX—XI пар), интернейронов и «-мотонейронов спинного мозга, синаптиче- ски связанных с аксонами пирамидных клеток. Экстрапир амидная система включает двигательную кору и совокупность ядер головного мозга (базальные ядра, черная субстанция, красное ядро, лью- исово тело, РФ, вестибулярные ядра и мозжечок), с помощью которых она управляет нейронами спинного мозга. Межполушарные функциональные связи осуществляются через мозолистое тело, а также комиссуры (спайки). Эти связи играют важную роль в деятельности организма, что показано в эксперименте на животных с рассечением мозолистого тел, а также подтверждаются результатами подобной операции у пациентов по медицинским показаниям (чтобы прекратить межполушарное распространение эпилептических судорог). При этом возникают различные нарушения: двигательные, речевые, узнавания с помощью зрительного и тактильного анализаторов. Например, больные с такими нарушениями могут рассказать о соматосенсорных стимулах, нанесенных на правую сторону тела, но не на левую, так как информация, поступающая в правые сенсомоторные области коры (от левой половины тела), не достигает речевых центров (они расположены в левом полушарии). Однако полушария могут выполнять отдельные функции независимо друг от друга. У здорового организма мозг работает как единое целое. 6.23. Методы исследования центральной нервной системы Наибольшее распространение получили методы регистрации биоэлектрической активности отдельных нейронов, суммарной активности нейронного пула или головного мозга в целом (электроэнцефалография), компьютерная томография (позитронно-эмиссионная томография, магнитно-резонансная томография) и др. Электроэнцефалография — это регистрация с поверхности кожи головы или с поверхности коры (последнее — в эксперименте) суммарного электрического поля нейронов мозга при их возбуждении (рис. 6.23). Происхождение волн ЭЭГ изучено недостаточно. Полагают, что ЭЭГ отражает ЛП множества нейронов — ВПСП, ТПСП, следовые — гиперполяризацию и деполяризацию, способные к алгебраической, пространственной и временной суммации. Эта точка зрения является общепризнанной, при этом участие ПД в формировании ЭЭГ отрицается. Так, например, W. Willes (2004) пишет: «Что касается потенциалов действия, то возникающие их ионные токи слишком слабы, быстры j50 мкВ 2 -
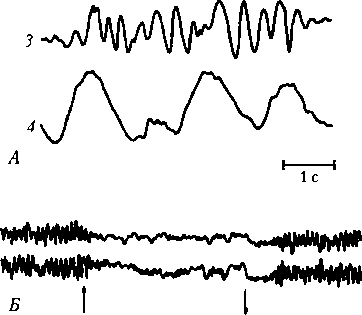
Рис. 6.23. Ритмы электроэнцефалограммы:
А — основные ритмы: 1 — а-ритм, 2 — 0-ритм, 3 — 9-ритм, 4 — 5-ритм; Б — реакция десинхронизации ЭЭГ затылочной области коры большого мозга при открывании глаз (?) и восстановление a-ритма при закрывании глаз (?) и несинхронизированны, чтобы их можно было зарегистрировать в виде ЭЭГ». Однако утверждение W. Willes не подкреплено экспериментальными фактами. Для его доказательства необходимо предотвратить возникновение ПД всех нейронов ЦНС и регистрировать ЭЭГ в условиях возникновения только ВПСП и ТПСП. Но это невозможно. Кроме того, в натуральных условиях ВПСП обычно являются начальной частью ПД, поэтому утверждать, что последние не участвуют в формировании ЭЭГ, оснований нет. Относительно слабости ионных токов при возникновении ПД следует заметить, что они формируют ПД в несколько раз большие по амплитуде, нежели ВПСП, а ЭЭГ — это регистрация суммарного электрического поля нейронов мозга, как, например, регистрация электрокардиограммы (ЭКГ). Утверждение о том, что ПД не участвуют в формировании ЭЭГ, так как они быстры и несинхронизированны, также не имеет оснований. По подсчетам, основанным на современных данных, головной мозг содержит около 100 млрд нейронов. Действительно, все они возбуждаются в разное время, но поскольку их огромное количество, то, конечно, многие из них возбуждаются синхронно. Например, a-ритм ЭЭГ в среднем составляет 10 ГЦ (10 колебаний в 1 с). Это означает, что период колебаний a-ритма составляет 0,1 с. Если разделить 100 млрд нейронов на 10, то получится, что в течение каждой 0,1 с могут возбуждаться до 10 млрд нейронов (ПД), причем каждый из них по несколько раз, т.е. имеются все условия для синхронизации возбуждений и обеспечения суммации электрических полей ПД в ритме ЭЭГ. Важно отметить что часть ВПСП и ТПСП нивелирует друг друга, часть ВПСП является составляющей ПД и только часть ВПСП, не достигающих величины КП, участвует в формировании ЭЭГ. Таким образом, ЭЭГ — это регистрация суммарного электрического поля ПД, ВПСП, ТПСП, следовых гиперполяризации и деполяризации нейронов. На ЭЭГ регистрируется четыре основных физиологических ритма: а-, 0-, 0- и 5-ритмы, частота и амплитуда которых отражают степень активности ЦНС. Метод вызванных потенциалов (ВП) заключается в регистрации изменений электрической активности мозга (электрического поля) (рис. 6.24), возникающих в ответ на раздражение сенсорных рецепторов, (обычный вариант). Позитронно-эмиссионная томография — метод функционального изотопного картирования мозга, основанный на введении в кровоток изотопов (13N, 18F, 15О) в соединении с дезоксиглюкозой. Чем активнее участок мозга, тем он больше поглощает меченой глюкозы. Радиоактивное излучение последней регистрируется специальными детекторами. Информация от детекторов поступает на компьютер, который создает «срезы» мозга на регистрируемом
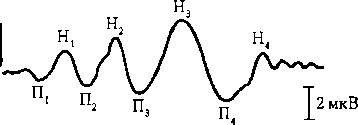
Рис. 6.24. Вызванные потенциалы у человека на вспышку света:
П — позитивные, Н — негативные компоненты ВП; цифровые индексы означают порядок следования позитивных и негативных компонентов в составе ВП. Начало записи совпадает с моментом включения вспышки света (стрелка) уровне, отражающие неравномерность распределения изотопа в связи с метаболической активностью мозговых структур, что позволяет судить о возможных поражениях ЦНС. Магнитно-резонансная томография позволяет выявить активно работающие участки мозга. Методика основана на том, что после диссоциации оксигемоглобина гемоглобин приобретает парамагнитные свойства. Чем выше метаболическая активность мозга, тем больше объемный и линейный кровоток в данном участке мозга и тем меньше отношение парамагнитного дезоксигемоглобина к оксигемоглобину. В мозге существует много очагов активации, что отражается в неоднородности магнитного поля. Стереотаксический метод. Метод позволяет вводить макро- и микроэлектроды, термопару в различные структуры головного мозга. Координаты структур мозга приведены в стереотаксических атласах. Посредством введенных электродов можно регистрировать биоэлектрическую активность данной структуры, раздражать или разрушать ее; через микроканюли можно вводить химические вещества в нервные центры или желудочки мозга; с помощью микроэлектродов (их диаметр менее 1 мкм), подведенных вплотную к клетке, можно регистрировать импульсную активность отдельных нейронов и судить об участии последних в рефлекторных, регуляторных и поведенческих реакциях, а также о возможных патологических процессах и применении соответствующих лечебных воздействий фармакологическими препаратами.
Вегетативная нервная система — это часть нервной системы, регулирующая работу внутренних органов, просвет сосудов, обмен веществ и энергии, обеспечивающих гомеостазис. 7.1. Общая характеристика вегетативной нервной системы Отделы ВНС В настоящее время общепризнанными являются два отдела ВНС: симпатический и парасимпатический. Влияния их на органы, как правило, противоположны (табл. 7.1); подробнее см. п. 7.2 и 7.3. В последние* годы получены убедительные факты, доказывающие наличие серотонинергических нервных волокон, идущих в составе симпатических стволов и усиливающих сокращения гладких мышц ЖКТ. Эти волокна являются пре- ганглионарными, они образуют синапсы с серотонинергическими нейронами вегетативных ганглиев, активация которых вызывает мощное усиление сокращений желудка и кишки, и формируют, по-видимому, серотонинергический отдел ВНС. Предлагается особым отделом ВНС считать интраорганную нервную систему и называть ее «метасимпатическая нервная система» — метасимпатическая часть (А. Д. Ноз- драчев). Блуждающий же нерв, согласно этому представлению, не имеет собственного эффекторного нейрона и рассматривается лишь как проводник ЦНС к эффекторным аппаратам метасимпатической нервной системы (греч. meta — следование за чем-либо, переход к чему-либо другому). Но это деление не по медиаторному признаку, а по месту расположения структурных элементов ВНС, поэтому целесообразно сохранить прежнее название — интра- органная нервная система. Кроме холинергических нейронов, в интраорганной нервной системе имеются адренергические, серотонинергические, по-видимому, пуринергические и др. Поскольку большинство постганглионарных симпатических волокон заканчивается непосредственно на эффекторных клетках, термин «метасимпатическая нервная система» с этим фактом не согласуется. Медиатор из сосудистых симпатических сплетений диффундирует к прилежащим клеткам. Эффекты раздражения блуждающего нерва устраняются блокадой М- и N-холинорецепто- ров, другие же нейроны интраорганной нервной системы при этом продолжают функционировать. Поэтому лишать главный парасимпатический нерв (блуждающий) собственного нейрона нет оснований, тем более что наличие собственного эффекторного холинергического нейрона у других парасимпатических нервов, иннервирующих органы головы и таза, не оспаривается. Таким образом, обсуждать следует только один вопрос: целесообразно ли называть Таблица 7.1 Реакции эффекторных органов на сигналы от автономных (вегетативных) нервов
Продолжение табл. 7.1
Окончание табл. 7. 1
1 Прочерк означает, что функциональная иннервация органа не обнаружена. 2 Знак «+» (от одного до трех) указывает, насколько важна активность адренергических и холинергических нервов в регуляции конкретных органов и функций. 3 In situ преобладает расширение, обусловленное метаболической авторегуляцией. 4 Физиологическая роль холинергической вазодилатации в указанных органах спорна. 5 В диапазоне физиологических концентраций адреналина, циркулирующего в крови, у сосудов скелетной мускулатуры и печени преобладает опосредуемая 0-рецепторами реакция расширения, а у сосудов других органов брюшной полости — реакция сужения, опосредуемая а-рецепторами. В сосудах почек и брыжейки есть, кроме того, специфические дофаминовые рецепторы, опосредующие расширение, которое, однако, не играет роли в большинстве физиологических реакций. 6 Холинергическая симпатическая система вызывает вазодилатацию в скелетной мускулатуре, но этот эффект не участвует в большинстве физиологических реакций. 7 Существует предположение, что адренергическими нервами снабжаются тормозные 0-ре- цепторы в гладких мышцах и тормозные а-рецепторы на парасимпатических холинергических (возбуждающих) ганглионарных нейронах ауэрбахова сплетения. 8 В зависимости от фазы менструального цикла, от концентрации в крови эстрогена и прогестерона, а также от других факторов. 9 Потовые железы ладоней и некоторых других областей тела («адренергическое потоотделение»). 10 Типы рецепторов, опосредующих определенные метаболические ответы, существенно варьируются у животных разных видов. 11 Парасимпатическая система вызывает вазодилатацию в слюнных железах посредством выделения совместно с ацетилхолином медиатора ВИП.
интраорганную нервную систему «метасимпа- тической»? По-видимому, нет, поскольку такие термины, как «центральная нервная система», «периферическая нервная система», ее часть — «интраорганная нервная система», даны не по медиаторному признаку, а по локализации. Исторически выделение симпатической и парасимпатической нервной системы тоже было осуществлено по анатомическому признаку. Однако, после открытия специфических медиаторов эффекторных нейронов этих отделов ВНС (Леви О., 1921), парасимпатические влияния стали оценивать как холинергические, а симпатические — как адренергические. Таким образом, симпатическая и парасимпатическая нервная системы имеют свои эффекторные медиаторы, их влияния строго специфичны и обычно разнонаправлены. Именно поэтому предлагавшийся ранее термин «ортосимпати- ческая нервная система» не был принят вместо термина «интраорганная нервная система» (Ле- ньел-Лавастин, 1924; Ботар, 1938). Дуга вегетативного рефлекса имеет те же звенья, что и дуга соматического рефлекса (рис. 7.1). Но имеются особенности ее организации: 1. Главное отличие заключается в том, что рефлекторная дуга ВНС может замыкаться вне ЦНС — интра- или электроорганно. 2. Афферентное звено дуги вегетативного рефлекса может быть образовано как собственными — вегетативными,
Рис. 7.1. Рефлекторная дуга вегетативного рефлекса:
1 — рецептор; 2 — афферентное звено; 3 — центральное звено; 4 — эфферентное звено; 5 — эффектор так и соматическими афферентными волокнами. 3. В дуге вегетативного рефлекса слабее выражена сегментированность, что повышает надежность вегетативной иннервации. Классификация вегетативных рефлексов (по структурно-функциональной организации):
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 225; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.93.221 (0.1 с.) |