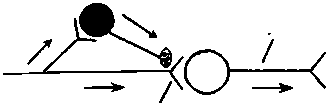Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
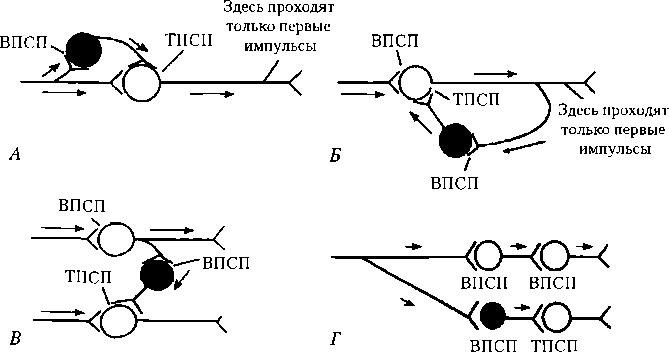
Имеется несколько разновидностей постсинаптического торможения.
Возвратное постсинаптическое торможение — тормоз Параллельное постсинаптическое торможение выполняет такую же роль — возбуждение блокирует само себя за счет распространения по коллатералям с включением тормозных клеток на своем пути и поступлением импульсов от тормозных клеток к нейрону-мишени (см. рис. 6.8, Б). В данном случае первый залп импульсов активирует постсинаптический нейрон, а следующий блокируется.
Прямым постсинаптическим торможением является, например, реципрокное торможение (от лат. reciprocus — взаимный). Так, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс: центр сгибания возбужден, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку — к центру мышцы-антагониста (разгибателю), что предотвращает ее сокращение (см. рис. 6.8, Г). Если бы возбуждались одновременно центры мышц сгибателей и разгибателей, сгибание конечности в суставе было бы невозможным. При возбуждении центра мышцы-разгибателя угнетается центр мышцы-сгибателя. В реципрокных взаимоотношениях находятся центры дыхания и глотания, вдоха и выдоха.
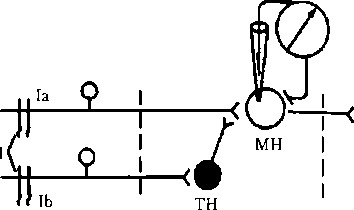
6.7. Пресинаптическое торможение Открытие (рис. 6.9). Известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных а-мотоней- ронов (а-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых а-мотонейронов. При этом мембранный потенциал и возбудимость исследуемых а-мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. Поскольку в опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги, было ясно, что они не возбуждаются вследствие процессов, происходящих в пресинаптическом
Рис. 6.9. Схема опыта, доказывающего наличие пресинаптического торможения. При раздражении 1а в МН регистрируется ПД, при опережающем раздражении на 20 мс lb в МН ПД не возникает:
1а — афферентный путь от мышечных рецепторов мышц-разгибателей (моносинаптическая рефлекторная дуга); 1Ь — афферентный путь от сухожильных рецепторов мышц-сгибателей; 1 — раздражающие электроды; МН — мотонейрон спинного мозга; TH — тормозной нейрон окончании. Это и определяет название данного вида торможения, так как возбуждение блокируется на подступах к постсинаптическому нейрону. В основе механизма пресинаптического торможения лежит деполяризация пресинаптических окончаний (тормозная пресинаптическая деполяризация — ТПД). В очаге деполяризации нарушается процесс распространения возбуждения, и, следовательно, поступающие к нервному окончанию импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуды, не обеспечивают выделения медиатора из пресинаптического окончания в синаптическую щель в достаточном количестве — постсинаптический нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные
Здесь проходят только первые импульсы
ТПД
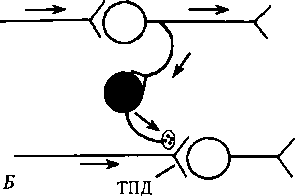
Рис. 6.10. Разновидности пресинаптического торможения:
А — параллельное; Б — латеральное. Нейроны: светлые — возбуждающие, черные — тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона-мишени (рис. 6.10). Снижение ПД в пресинаптической терминали (частичная деполяризация) всего лишь на 5 % уменьшает ВПСП на постсинаптической мембране на 50 %. Торможение (деполяризация) терминали после одного афферентного залпа продолжается 300—400 мс; медиатором служит ГАМК, которая действует на TAMKj-рецепторы. ТПД является следствием повышения проницаемости для С1~ и выхода его из клетки. Этот факт свидетельствует о том, что в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий первичный транспорт С1_ внутрь клетки вопреки электрическому градиенту. Под действием ГАМК тормозных нейронов и последующего повышения проницаемости мембраны для С1_ ионы последнего начинают выходить наружу согласно электрическому градиенту, но вопреки концентрационному. Это приводит к деполяризации пресинаптических терминалей и ухудшению их способности проводить импульсы к постсинаптическому нейрону. Роль ГАМК2-рецепторов на пресинаптических окончаниях изучена недостаточно. Хотя известно, что активация ГАМК2- рецепторов на постсинаптической мембране нейрона ведет к выходу ионов К+ из клетки (гиперполяризация) и снижению возбудимости. Известны параллельное и латеральное пресинаптическое торможение (см. рис. 6.10). Возвратное пресинаптическое торможение на уровне спинного мозга (по типу возвратного постсинаптического торможения) у млекопитающих обнаружить не удалось, хотя у лягушек оно выявлено. В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рис. 6.8 и 6.10. Тем не менее все варианты пре- и постсинаптического торможения можно объединить в две группы: 1) когда распространяющееся возбуждение блокирует собственный путь с помощью вставочных тормозных клеток (параллельное и возвратное торможение) и 2) когда распространяющееся возбуждение блокирует другие нервные пути с помощью включения тормозных клеток (латеральное и прямое торможение). Поскольку тормозные клетки сами могут быть заторможены другими тормозными нейронами, это может облегчить распространение возбуждения, т.е. наблюдается феномен растормаживания. Иногда в качестве разновидности центрального торможения выделяют торможение вслед за возбуждением. Исходя из имеющихся фактов, особым механизмом торможения считать его нельзя, поскольку оно может быть результатом следовой гиперполяризации нейронов либо возвратного торможения. В спинном мозге это осуществляется с помощью клеток Реншоу. Песси- мальное торможение (пессимум Введенского), наблюдаемое в эксперименте на нервно-мышечном препарате, в ЦНС в физиологических условиях также, по-видимому, не встречается. «Общее торможение» (И. С. Беритов) тоже не является каким-то особым механизмом торможения. В условиях натуральной жизнедеятельности организма это первая фаза ориентировочного рефлекса — прекращение текущей деятельности с фиксацией позы, превентивное торможение; по П. В. Симонову, это феномен, но не механизм. 6.8. Общая характеристика торможения в центральной нервной системе
|
||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 247; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.43.140 (0.007 с.) |