Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные медиаторы центральной нервной системы
Амины (дофамин, норадреналин, серотонин, гистамин) встречаются в разных отделах ЦНС, в значительных количествах — в нейронах ствола мозга. Амины обеспечивают возникновение процессов возбуждения и торможения, например, в промежуточном мозге, в черной субстанции, в лимбической системе, в полосатом теле. Серотонин является возбуждающим и тормозным медиатором в нейронах ствола мозга, тормозным — в коре большого мозга. Известно семь типов серотонино- рецепторов (5-НТ, S-рецепторы), большинство из них метаботропные (вторые посредники — цАМФ и ИФ3/ДАГ). Ионотропным является 83-рецептор (имеется, в частности, в ганглиях ВНС). Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций. Особенно много его в ядрах шва (ЯШ), лимбической системе. Аксоны этих нейронов проходят в бульбоспинальных путях и оканчиваются на нейронах различных сегментов спинного мозга. Здесь они контактируют с клетками преганглио- нарных симпатических нейронов и со вставочными нейронами желатинозной субстанции. Полагают, что часть этих симпатических нейронов (а может быть, и все) являются серотонинергическими нейронами ВНС. Их аксоны, согласно последним данным, идут к органам ЖКТ и оказывают мощное стимулирующее влияние на его моторику. Повышение уровня серотонина и норадреналина в нейронах ЦНС типично для маниакальных состояний, снижение — для депрессивных. Норадреналин является возбуждающим медиатором в гипоталамусе, в ядрах эпиталамуса, тормозным — в клетках Пуркинье мозжечка. В ретикулярной формации (РФ) ствола мозга и гипоталамусе обнаружены а- и 0-адре- норецепторы. Норадренергические нейроны сконцентрированы в области голубого пятна (средний мозг), где их насчитывается всего несколько сотен, но ответвления их аксонов встречаются по всей ЦНС. Дофамин является медиатором нейронов среднего мозга, гипоталамуса. Дофаминорецепторы подразделяют на Дг и Д2-подтипы. Д]-рецепторы локализуются на клетках полосатого тела, действуют посредством дофаминчувствительной аденилатциклазы, как и Д2-рецепторы. Последние обнаружены в гипофизе. При действии на них дофамина угнетаются синтез и секреция пролактина, окситоцина, меланоцитстимулирующе- го гормона, эндорфина. Д2-рецепторы найдены на нейронах полосатого тела, где их функция пока не очень ясна. Содержание дофамина в нейронах ЦНС повышено при шизофрении и снижено при паркинсонизме.
Гистамин реализует свое влияние с помощью вторых посредников (цАМФ и ИФз/ДАГ). В значительной концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса — здесь же локализовано основное количество ги- стаминергических нейронов. В остальных отделах ЦНС уровень гистамина очень низок. Медиаторная роль гистамина изучена мало. Выделяют Н,-, Н2- и Н 3-гистаминорецепторы. Н,-рецепторы имеются в гипоталамусе и участвуют в регуляции потребления пищи, в терморегуляции, секреции пролактина и антидиуретического гормона (АДГ). Н2-рецепторы обнаружены на глиальных клетках. Ацетилхолин встречается в коре большого мозга, в спинном мозге. Известен в основном как возбуждающий медиатор; в частности, является медиатором а-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина а-мотонейро- ны по коллатералям своих аксонов передают возбуждающее влияние на тормозные клетки Реншоу; ацетилхолин имеется в РФ ствола мозга, в гипоталамусе. Обнаружены М- и N-холинорецепторы. Установлено семь типов М-холиноре- цепторов; основными являются и М2-рецепторы. Мх-холинорецепторы локализуются на нейронах гиппокампа, полосатого тела, коры большого мозга, М2-холинорецепторы — на клетках мозжечка, ствола мозга. N-холинорецепторы довольно плотно расположены в области гипоталамуса и покрышки. Эти рецепторы изучены достаточно хорошо, они выделены с помощью «-бунгаротоксина (основной компонент яда ленточного крайта) и а-нейротоксина, содержащегося в яде кобры. При взаимодействии ацетилхолина с N-холинорецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. При взаимодействии ацетилхолина с М-холинорецептором активация ионных каналов (К+, Са2+) осуществляется с помощью вторых внутриклеточных посредников (цАМФ — циклический аденозинмонофосфат — для М2-рецептора; ИФ3/ДАГ — для Mj-рецептора).
Ацетилхолин активирует и тормозные нейроны с помощью М-холинорецеп- торов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре. Аминокислоты. Глицин и у-ами- номасляная кислота (ГАМК) являются тормозными медиаторами в синапсах ЦНС и действуют на соответствующие рецепторы, глицин — в основном, в спинном мозге, ГАМК — в коре большого мозга, мозжечке, стволе мозга, спинном мозге. Передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы а-глутамат и а-аспартат. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Глутамат — это основной возбуждающий медиатор ЦНС (75 % возбуждающих синапсов мозга). Глутамат реализует свое влияние посредством метаботропных (связанных с активацией цАМФ и ИФуДАГ) и ионотропных (связанных с К+-, Са2+-, Ма+-ионными каналами рецепторов). Полипептиды встречаются в синапсах различных отделов ЦНС. Энкефалины и эндорфины — опиоидные медиаторы нейронов, блокирующих, например, болевую импульсацию. Реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы; много их также на клетках черной субстанции, ядрах промежуточного мозга и солитарного тракта, имеются и на клетках голубого пятна, спинного мозга. Их лигандами являются Р-эндорфин, динорфин, лей- и метэнкефалины. Различные опиатные рецепторы обозначаются буквами греческого алфавита: а, е, к, ц, %. Вещество Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга. Это позволило предположить, что вещество Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочные нейроны. Большое количество вещества Р содержится в гипоталамической области. Различают два вида рецепторов вещества Р: рецепторы типа SP-E (Pj), расположенные на нейронах коры большого мозга, и рецепторы типа SP-P (Р2), расположенные на нейронах мозговой перегородки. Вазоинтестинальный пептид (ВИП), соматостатин, холецистокинин (ХЦК) также выполняют медиаторную функцию. ВИП-рецепторы и рецепторы к соматостатину выявлены на нейронах головного мозга. Рецепторы к ХЦК обнаружены на клетках коры большого мозга, хвостатого ядра, обонятельных луковиц. Действие ХЦК на рецепторы повышает проницаемость мембран для Са2+ посредством активации аденилатциклазной системы. Ангиотензин участвует в передаче информации о потребности организма в воде. Рецепторы к ангиотензину обнаружены на нейронах коры большого мозга, среднего и промежуточного мозга. Связывание ангиотензина с рецепторами вызывает увеличение проницаемости клеточных мембран для Са2+. Эта реакция обусловлена процессами фосфорилирования мембранных белков вследствие активации аденилатциклазной системы и изменением синтеза простагландинов. Люлиберин участвует в формировании половой потребности. Пурины (АТФ, аденозин, АДФ) выполняют в основном моделирующую функцию. В частности, АТФ в спинном мозге выделяется вместе с ГАМК. Рецепторы к АТФ весьма разнообразны: Одни из них ионотропные, другие — метаботропные. АТФ и аденозин ограничивают перевозбуждение ЦНС и участвуют в формировании болевых ощущений.
Гипоталамические нейрогормоны, регулирующие функцию гипофиза, также выполняют медиаторную роль. Физиологические эффекты действия некоторых медиаторов головного мозга. Дофамин участвует в формировании чувства удовольствия, в регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений. Серотонин ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание. Эндорфины, энкефалины, пептид, вызывающий 5-сон, дают анти- болевые эффекты, повышают устойчивость к стрессу, способствуют сну. Простагландины вызывают повышение свертываемости крови, изменение тонуса гладких мышц, усиливают физиологические эффекты медиаторов и гормонов. Олигопептиды — медиаторы настроения, полового поведения, передачи ноцицептивного возбуждения от периферии к ЦНС, формирования болевых ощущений. В последние годы получены факты, вызвавшие необходимость внесения коррективов в известный принцип Дейла. Так, согласно принципу Дейла, один нейрон синтезирует и использует один и тот же медиатор во всех разветвлениях своего аксона («один нейрон — один медиатор»). Однако выяснилось, что, кроме основного медиатора, в окончаниях аксона могут выделяться и другие, сопутствующие медиаторы (комедиаторы), играющие модулирующую роль или более медленно действующие. Кроме того, в тормозных нейронах в спинном мозге в большинстве случаев имеется два быстродействующих типичных медиатора в одном тормозном нейроне — ГАМК и глицин. Поэтому принцип Дейла в новой редакции сначала звучал так: «Один нейрон — один быстрый медиатор», а затем: «Один нейрон — один быстрый синаптический эффект». Не исключено, что принцип Дейла вообще будет представлять лишь исторический интерес. Таким образом, нейроны ЦНС возбуждаются или тормозятся в основном под влиянием специфических медиаторов. Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны и вторых посредников. Это явление особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и в периферических синапсах организма. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — только торможение, в синапсах гладкой мускулатуры ЖКТ — только возбуждение. Катехоламины тормозят сокращения желудка и кишечника, но стимулируют сердечную деятельность. Глутамат является только возбуждающим медиатором ЦНС.
6.3. Механизм возбуждения нейронов При возбуждении нейронов потребление кислорода возрастает. Источником энергии служит в основном глюкоза крови, собственных небольших запасов гликогена хватает лишь на 3-5 мин работы нейрона. Передача сигнала в химических синапсах ЦНС подобна таковой в нервно-мышечном синапсе, однако имеет ряд отличительных особенностей, основные из которых приводятся ниже. Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных импульсов и их взаимодействие. Одна везикула (квант медиатора) содержит 1—10 тыс. молекул медиатора. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 200—300 квантов медиатора; при этом возникает небольшой ВПСП — около 0,05 мВ (миниатюрный ВПСП). Необходимо учесть, что одновременно могут возникать не только возбуждающие, но и тормозные потенциалы. Пороговый потенциал нейрона равен 5—10 мВ, поэтому ясно, что для возбуждения нейрона требуется некоторое множество импульсов. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль Са2+, причем количество медиатора прямо пропорционально входу ионов Са2+, при этом четыре иона Са2+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени по различным входам в результате пространственной суммации ВПСП возникает ПД нейрона-мишени. ВПСП возникает вследствие суммарного тока в клетку и из клетки через ионные каналы различных ионов согласно электрохимическому градиенту. Поступивший в пресинаптическое окончание Са2+ удаляется в среду с помощью Са-насоса. Прекращение действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично — с помощью разрушения специальными ферментами. Например, норадреналин расщепляется МАО и катехолметилтрансфе- разой, ацетилхолин гидролизуется аце- тилхолинтрансферазой, имеющейся в синаптической щели и встроенной в постсинаптическую мембрану. Прекращение действия избытков медиатора на постсинаптическую мембрану предотвращает десенситизацию — снижение чувствительности постсинаптической мембраны к действующему медиатору. В возникновении ПД в нейронах принимают участие ионы Са2+, ток которых в клетку более медленный, чем ток Na+, играющий главную роль в формировании ПД-нейронов. В частности, в дендритах клеток Пуркинье мозжечка выявлены не только быстрые натриевые потенциалы, но и медленные кальциевые. В телах некоторых нервных клеток потенциал действия создается преимущественно за счет Са2+.
Место возникновения ВПСП, вызывающих ПД нейрона, — тело нейрона, поскольку постсинаптические мембраны этих синапсов располагаются в непосредственной близости от аксонного холмика, где начинается развитие ПД. Близость ВПСП, возникающих в теле нейрона, к аксонному холмику обеспечивает их участие в механизмах генерации ПД. Однако подавляющее большинство синапсов (в частности, в коре большого мозга, согласно расчетам, 98 %) находится на дендритах. Площадь мембраны тела нейронов занята синапсами на 40 %, дендритов — на 75 %. Синапсов в коре большого мозга в 40 тыс. раз больше, чем самих нейронов. Место возникновения ПД нейрона. Формирование ПД начинается на мембране аксонного холмика (генераторный пункт нейрона). Синапсы на нем отсутствуют, возбудимость мембраны аксонного холмика в 3—4 раза превосходит возбудимость мембраны тела нейрона, что объясняется более высокой (примерно в 7 раз) концентрацией Na-каналов на аксонном холмике (на нем отсутствует миелиновая оболочка). ВПСП, возникающие в любых участках сомы нейрона суммируются, их суммарное электрическое поле достигает аксонного холмика и вызывает его деполяризацию до КП, в результате чего начинает развиваться ПД на аксонном холмике. Это связано с тем, что постоянная длины мембраны тела нейрона (расстояние, на котором исходная амплитуда ВПСП уменьшается на 37 %) составляет 1-2 мм, а диаметр тела нейрона — всего лишь 10-80 мкм. Поэтому электрическое поле начального возбуждения аксонного холмика обеспечивает деполяризацию тела нейрона до КП, в результате чего возникает ПД нейрона, который проводится по аксону к другой клетке. На восходящей части ПД нейрона регистрируется перегиб на уровне 30— 40 мВ (задержка в развитии ПД), вследствие того что возбудимость мембраны тела нейрона в 3—4 раза ниже возбудимости мембраны аксонного холмика, что также является подтверждением
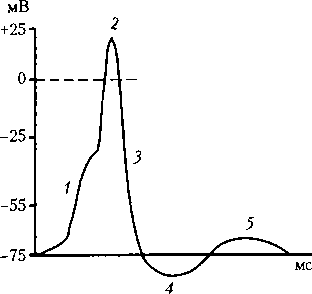
Рис. 6.2. Типичный ПД нейронов ЦНС: 1 — деполяризация; 2 — инверсия; 3 — реполяризация; 4 — следовая гиперполяриэация; 5 — следовая деполяризация
описанного механизма формирования ПД нейрона (рис. 6.2). Таким образом, ПД нейрона — это единый одновременный процесс возбуждения аксонного холмика и тела нейрона. Роль дендритов в возникновении возбуждения нейрона. Дендритные синапсы называют модуляторами. Это связано с тем, что они удалены на значительное расстояние от генераторного пункта нейрона — аксонного холмика. Поэтому суммарное электрическое поле их ВПСП не может вызвать достаточную деполяризацию аксонного холмика и обеспечить возникновение ПД, а определяет лишь возбудимость нейрона.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 109; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.119.126.80 (0.022 с.) |