
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности нервного и гуморального механизмов регуляции функций организма
Нервная регуляция является ведущей — ей подчиняется эндокринная система. Различная скорость связи: нервный импульс распространяется с огромной скоростью — до 120 м/с. Вещество, попадающее в кровь проходит один раз через весь организм (большой и малый круг кровообращения) за 22 с. У гуморального и нервного механизмов регуляции функций способы связи различны: у нервной системы — нервный импульс как универсальный сигнал, а у гуморального механизма связь с регулируемым органом или тканью осуществляется с помощью различных химических веществ (гормоны, медиаторы, метаболиты и парагормоны). Некоторые медиаторы, например катехоламины, попадая в кровь, могут действовать не только в месте их выделения нервными окончаниями, но и на другие органы и ткани организма. Нервная система может оказывать точное, локальное влияние на отдельный орган или даже на группу клеток этого органа (различная точность связи). Так, нервная система может вызывать сокращения мышц одного какого-то пальца руки, не вызывая сокращений мышц всей конечности или других пальцев. Химические вещества, попадая в кровь, разносятся по всему организму и действуют нередко на многие органы и ткани — системный (генерализованный) характер влияния. Например, адреналин, тироксин, попадая в кровь, разносятся по всему организму и действуют на клетки всех органов и тканей организма. Следует, однако, заметить, что и у гуморального механизма нередко имеется точный адресат воздействий. Так, кортикотропин хотя и разносится с кровью по всему организму, но действует только на кору надпочечников. Тиреотропный гормон (ТТГ) регулирует функцию щитовидной железы. В свою очередь, и нервная система может оказывать генерализованное влияние, что характерно для симпатической нервной системы — в экстремальных условиях она мобилизует ресурсы всего организма для достижения цели. Возможно противоположное влияние БАВ на один и тот же орган в зависимости от точки приложения действия этого химического вещества. Адреналин, например, при непосредственном действий на сердце стимулирует его работу, а при введении в ликвор, возбуждая центры блуждающих нервов, он, наоборот, тормозит ее. Угольная кислота, действуя прямо на кровеносные сосуды, вызывает их расширение, а посредством возбуждения центра кровообращения — сужение. Поэтому результат действия химического вещества может зависеть от того, проникает ли оно в цереброспинальную жидкость через гематоэнцефалический барьер (ГЭБ) или нет.
Функции ГЭБ. Гематоэнцефалический барьер выполняет регулирующую функцию, которая заключается в том, что он формирует особую внутреннюю среду мозга, обеспечивающую оптимальный режим деятельности нервных клеток, и избирательно пропускает многие гуморальные вещества, например углекислый газ. Последний, действуя на дыхательный, сердечно-сосудистый и другие центры, регулирует функцию соответствующих систем организма. Барьерную функцию выполняет эндотелий капилляров мозга, а также базальная мембрана, окружающая капилляр снаружи. Их эндотелий имеет очень мало пор, узкие щелевые контакты между клетками, почти лишен «окон». Составной частью являются также глиальные клетки, образующие своеобразные футляры вокруг капилляров, покрывающие около 90 % их поверхности. Наибольший вклад в развитие представлений о ГЭБ внесла Л. С. Штерн. Этот барьер пропускает воду, ионы, глюкозу, аминокислоты, газы, но задерживает многие физиологически активные вещества: дофамин, инсулин, адреналин, тироксин, серотонин. Однако в ГЭБ существуют «окна», через которые соответствующие клетки мозга (хеморецепторы) получают прямую информацию о наличии в крови гормонов и других, не проникающих через барьер веществ; клетки мозга выделяют и свои нейросекреты. Зоны мозга, не имеющие собственного ГЭБ, — это гипофиз, эпифиз, некоторые отделы гипоталамуса и продолговатого мозга. Защитная функция ГЭБ— это предотвращение попадания микробов, чужеродных или токсичных веществ экзо- и эндогенной природы в межклеточные пространства мозга. ГЭБ не пропускает многие лекарственные вещества, что необходимо учитывать в клинической практике — при воздействии на ЦНС следует подбирать препарат, который проникает через ГЭБ, иначе его действие не даст эффекта. 2.4. Системный принцип регуляции функций организма
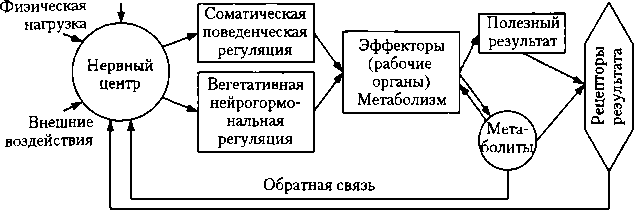
Понятие о системном принципе регуляции. Этот принцип регуляции заключается в том, что различные показатели организма поддерживаются на оптимальном уровне с помощью многих органов и систем. Например, Ро2 и Рсо2 обеспечиваются деятельностью сердечно-сосудистой, дыхательной, нервно- мышечной системы, системой крови. Они объединяются в различные функциональные системы. Представление о функциональных системах разработал П. К. Анохин в середине XX в. В последние годы теория функциональных систем успешно развивается К. В. Судаковым и его сотрудниками. Функциональная система — это динамическая совокупность различных
Различают гомеостатические и поведенческие функциональные системы. Гомеостатические системы обеспечивают поддержание на оптимальном для метаболизма уровне различных показателей организма (например, pH, АД). Это достигается посредством изменения интенсивности работы внутренних органов. Полезным результатом поведенческой функциональной системы является социальный, бытовой успех (получение образования, достижение выдающихся спортивных результатов и т.п.), удовлетворение биологической потребности. Поведенческие реакции нередко являются составным элементом гомеостатической функциональной системы (например, поиск и прием воды и пищи в случае их недостатка в организме). Поведенческая функциональная система может формироваться организмом с целью избавления или избегания действия вредных раздражителей. В свою очередь, любые поведенческие реакции организма обеспечиваются деятельностью функциональных гомеостатических систем. Таким образом, поведенческие функциональные системы включают в реакции внутренние органы и опорнодвигательный аппарат. Поведенческая функциональная система описана в п. 19.2. Функциональные системы формируются из физиологических систем. Физиологическая система — генетически сформированная совокупность органов и тканей организма, выполняющая определенную функцию (например, система дыхания). Структура физиологической системы включает: 1) управляющее устройство — нервный центр, представляющий собой объединение ядер различных уровней ЦНС; 2) выходные каналы (нервы и гормоны); 3) исполнительные органы — эффекторы, обеспечивающие в ходе физиологической деятельности поддержание регулируемого процесса, показателя на некотором оптимальном уровне (полезный результат деятельности функциональной системы); 4) рецепторы результата (сенсорные рецепторы) — датчики, воспринимающие информацию о параметрах отклонения регулируемого процесса, показателя от оптимального уровня; 5) канал обратной связи (входные каналы), информирующие нервный центр с помощью афферентной им- пульсации от рецепторов результата о достаточности или недостаточности эффекторных усилий по поддержанию регулируемого процесса, показателя на оптимальном уровне. Эти импульсы по каналам обратной связи поступают в нервный центр, регулирующий тот или иной показатель.
Обратная связь осуществляется также с помощью гуморальных веществ. Метаболиты, попадая в кровь, а с кровью — в ЦНС, действуют и на соответствующие центры, изменение активности которых оказывает корригирующее влияние на органы и ткани организма. Метаболиты действуют также на сам работающий орган, что ведет к расширению сосудов и улучшению обмена веществ в органе, и на рецепторы рабочего органа (или органов) — рецепторы результата, что тоже отражается на активности рецепторов и, естественно, на импульса- ции в афферентных путях, проводящих импульсы в ЦНС по принципу обратной связи и способствующие нормализации регулируемого показателя. Например, при увеличении АД крови в большей степени начинают раздражаться барорецепторы рефлексогенных сосудистых зон, в результате чего увеличивается поток импульсов в ЦНС — в центр кровообращения. Взаимодействие нейронов этого центра н изменения интенсивности эфферентной импульсации ведут к ослаблению деятельности сердца и расширению кровеносных сосудов, АД крови снижается. Если же одной физиологической системы оказалось недостаточно и показатель организма остается повышенным или пониженным (например, АД), то формируется функциональная система, включающая дополнительные физиологические системы — выделительную (больше воды выводится из организма, АД снижается — нормализуется), эндокринную (адреналин, ангиотензин). Основные принципы формирования и деятельности гомеостатических функциональных систем зрелого организма Изоморфизм функциональных систем. Все функциональные системы построены принципиально одинаково. Вместе с тем они могут отличаться друг от друга по степени разветвленности как центральных, так и периферических механизмов. Необходимо подчеркнуть, что системообразующим фактором является полезный для жизнедеятельности организма приспособительный результат. Ряд гомеостатических функциональных систем представлен исключительно внутренними генетически детерминированными механизмами вегетативной нервно-гормональной регуляции и не включает механизмы поведенческой соматической регуляции. Примером являются функциональные системы, определяющие оптимальные для обмена веществ кровяное давление, содержание ионов в крови, pH внутренней среды организма, не вызывающие ощущений и не побуждающие организм к поведенческим реакциям. Ряд гомеостатических функциональных систем включает целенаправленное поведение во внешней среде на базе доминирующих мотивационных возбуждений, отражающих сдвиги различных показателей метаболизма, сопровождающиеся возникновением соответствующих ощущений. В этом случае системообразующим фактором является также и мотивация. Примерами таких функциональных систем являются системы, обеспечивающие поддержание оптимального уровня питательных веществ в организме, осмотического давления и объема жидкости, возбуждение которых сопровождается возникновением соответствующих ощущений. В этом случае опорно-двигательный аппарат выступает как составная часть эффектора — рабочего органа. Достижение полезного результата обеспечивается усилением их сократительной деятельности скелетной мускулатуры, усилением деятельности сердца, стимуляцией дыхания, мобилизацией крови из депо организма.
Взаимосодействие функциональных систем осуществляется помуль- типараметрическому принципу, так как показатели внутренней среды организма являются результатом согласованной деятельности многих систем, и поэтому они взаимосвязаны. Изменение величины одного показателя может привести к изменению величины других показателей. Например, избыточное поступление воды в организм сопровождается увеличением объема циркулирующей крови (ОЦК), повышением АД, снижением осмотического давления плазмы крови. В функциональной системе, поддерживающей оптимальный уровень газового состава крови, одновременно осуществляется взаимодействие pH, Рсо2, Ро2. Изменение одного из этих параметров немедленно приводит к изменению количественных характеристик других параметров. На основе принципа мультипараметрического взаимосо- действия все функциональные системы гомеостатического уровня фактически объединяются в единую функциональную систему гомеостазиса, включающую и поведенческие реакции. Системообразующим фактором функциональной системы любого уровня является полезный для жизнедеятельности организма приспособительный результат, необходимый в данный момент, и доминирующая мотивация. Все множество полезных приспособительных результатов можно объединить в две группы: 1) поддержание постоянства внутренней среды организма; 2) достижение результата в социальной деятельности. В системоге- незе выделяют два периода: пренатальный (внутриутробный) и постнатальный (внеутробный). Особенности пренатального системо- генеза. Согласно П. К. Анохину, системо- генез — избирательное созревание и развитие функциональных систем в пре- и постнатальном онтогенезе. В отличие от понятия «морфогенез», предложенного А. Н. Северцевым (развитие органов в онтогенезе), термин «системогенез» отражает развитие в онтогенезе различных по функции и локализации структурных образований, которые объединяются в полноценную функциональную систему, обеспечивающую новорожденному выживание. В настоящее время термин «системогенез» применяется в более широком смысле, при этом под систе- могенезом понимают процессы не только онтогенетического созревания функциональных систем, но и формирование и преобразование функциональных систем в ходе жизнедеятельности зрелого организма.
Рассмотрим общие принципы формирования функциональных систем в онтогенезе по П. К. Анохину. Принцип минимального обеспечения реализуется с помощью гетерохронной закладки и гетерох- ронного созревания отдельных компонентов функциональной системы. Полное завершение развития функциональных систем организма наблюдается в ходе постнатального онтогенеза. Физиологические же системы детского организма отличаются от функциональных систем взрослого организма тем, что раньше созревают те фрагменты органа (системы), которые необходимы для формирования функциональной системы, обеспечивающей выживание организма (например, сосание). На основе принципа минимального обеспечения функциональная система начинает играть приспособительную роль задолго до того, как все элементы органа завершат свое структурное оформление. В онтогенезе созревают в первую очередь те функциональные системы, без которых невозможно дальнейшее развитие организма. Например, у плода развивается функциональная система, поддерживающая постоянство газового состава крови с помощью плацентарного кровообращения. К моменту рождения сформированы функциональные системы поддержания температуры тела, регуляции осмотического давления и др. Наиболее зрелой, хотя и не завершившей свое развитие к моменту рождения, является функциональная система поддержания постоянства газового состава крови. К моменту рождения все компоненты, вне зависимости от их количества, должны составить единую функциональную систему, так как только в таком случае она сможет обеспечить новорожденному выживание в конкретных условиях постнатального онтогенеза. Так, если у новорожденного т. orbi- kularius oris по причине незрелости не дает должной герметизации ротовой полости, функциональная система сосательного акта никакого положительного эффекта новорожденному не обеспечит из-за отсутствия обязательного звена в составе комплекса ее моторных эффекторов. Принцип минимального обеспечения реализуется также с помощью фрагментации органов в процессе пренатального онтогенеза. В пределах одного и того же органа прежде всего развиваются те фрагменты, которые обеспечивают к моменту рождения возможность функционирования некоторой целостной функциональной системы. Например, разные клеточные группы ядра лицевого нерва созревают с разной скоростью, причем с наибольшей скоростью дифференцируются те фрагменты ядра, которые в будущем должны обеспечить функциональную систему сосания; соответственно, и нервные волокна, идущие к сосательным мышцам, миелинизируются и образуют синаптические контакты раньше, чем нервные волокна, направляющиеся к лобным мышцам. Принцип консолидации компонентов функциональной системы — объединение в функциональную систему отдельных фрагментов, развивающихся в различных частях организма. Ведущую роль в этом процессе играет ЦНС. Например, сердце, сосуды, дыхательный аппарат, кровь объединяются в функциональную систему поддержания постоянства газового состава внутренней среды на основе совершенствования связей между различными отделами ЦНС, а также на основе развития нервных связей между ЦНС и соответствующими периферическими органами. Сравнительная характеристика физиологических и функциональных гомеостатических систем Главное сходство заключается в том, что обе системы включают одни и те же блоки: 1) управляющее устройство (нервный центр или совокупность нервных центров); 2) выходные каналы (нервы и гормоны); 3) исполнительные органы (эффекторы); 4) рецепторы результата; 5) каналы обратной связи. Главные отличия состоят в их происхождении и назначении — структура и функция физиологической системы генетически обусловлены, их число строго ограничено; структура функциональных систем является динамической, число физиологических систем, формирующих функциональную систему, различно; число самих функциональных систем различного уровня и сложности огромно. 2.5. Типы регуляции функций организма и их надежность Типы регуляции. Как отмечалось выше, имеется два типа регуляции — по отклонению и по опережению. Рассмотрим их подробнее. Регуляция по отклонению является ведущей в организме и заключается в том, что всякое отклонение от оптимального уровня регулируемого показателя мобилизует физиологические системы к восстановлению его на прежнем уровне. Регуляция по отклонению осуществляется с помощью системного комплекса — канала отрицательной обратной связи, обеспечивающего разнонаправленное влияние — усиление стимулирующих механизмов управления (в случае ослабления показателей процесса) или их ослабление (в случае чрезмерного увеличения показателей процесса). В отличие от отрицательной обратной связи, положительная обратная связь встречается в организме редко; примером ее является усиление начавшейся секреции пищеварительных соков в желудке после приема пищи с помощью продуктов гидролиза, всосавшихся в кровь. Положительная обратная связь оказывает только однонаправленное влияние на развитие процесса, находящегося под контролем управляющего комплекса. Поэтому положительная обратная связь делает систему неустойчивой, неспособной обеспечить стабильность регулируемого процесса в пределах физиологического оптимума. Например, если бы АД регулировалось по принципу положительной обратной связи, то в случае понижения АД действие регуляторных механизмов привело бы к еще большему его снижению, а в случае повышения — к еще большему его увеличению. Основные показатели внутренней среды поддерживаются в диапазоне колебаний, не нарушающих оптимальный ход жизнедеятельности организма. Из этого вытекает, что представление о показателях внутренней среды организма как о стабильных величинах относительно. Вместе с тем выделяют «жесткие» показатели, которые поддерживаются соответствующими функциональными системами на сравнительно фиксированном уровне и отклонение которых от этого уровня оказывается минимальным, так как чревато серьезными нарушениями метаболизма. Примерами «жестких» показателей являются уровень осмотического давления, величина pH. Выделяют также «пластичные» («мягкие») показатели, отклонение которых от оптимального уровня допускается в широком физиологическом диапазоне. Примерами «пластичных» показателей являются концентрация питательных веществ крови, величина кровяного давления. Регуляция по опережению встречается реже в организме. При этом регулирующие механизмы включаются до реального изменения параметра регулируемого процесса (показателя) на основе информации, поступающей в нервный центр, о возможном изменении регулируемого процесса (показателя) в будущем. Например, усиление вентиляции легких при физической нагрузке начинается раньше изменений газового состава крови человека. Это происходит благодаря афферентной импульсации от проприорецепторов активно работающих мышц к моторным центрам и взаимодействию их с дыхательным центром. Терморецепторы (детекторы температуры), находящиеся внутри тела, обеспечивают контроль за температурой внутренних областей тела. Терморецепторы кожи, в основном, играют роль детекторов температуры окружающей среды (возмущающий фактор). При значительных отклонениях температуры окружающей среды создаются предпосылки возможного изменения температуры внутренней среды организма. Однако в норме этого не происходит, так как импульсация от терморецепторов кожи, непрерывно поступая в гипоталамический терморегуляторный центр, позволяет ему заранее изменить направленность терморегуляторных процессов. Механизм условного рефлекса также обеспечивает регуляцию по опережению. Например, деятельность пищеварительной системы стимулируется при виде, запахе пищи и даже при звоне столовой посуды. Регуляция по опережению дополняет регуляцию по отклонению и делает единую функциональную систему — организм — весьма устойчивой в различных условиях жизнедеятельности.
3.1. История. Сущность процесса возбуждения История. В 1786 г. профессор анатомии Болонского университета Луиджи Гальвани выполнил два опыта на препаратах задних лапок лягушек со снятой кожей. Впервом опыте, подвешивая препарат с помощью медного крючка к железным перилам балкона, он наблюдал сокращение мышц, когда те касались перил балкона. Л. Гальвани предположил, что сокращение мышц возникало как следствие воздействия на них электричества, источником которого являются «животные ткани» — мышцы и нервы. Однако другой итальянский исследователь — физик и физиолог А. Вольта оспорил гипотезу, выдвинутую Л. Гальвани. По его мнению, причиной сокращения мышц был электрический ток, возникающий в области контакта двух разнородных металлов (медь и железо — гальваническая пара) с тканями лягушки. Чтобы проверить справедливость своей гипотезы, Л. Гальвани поставил второй опыт, в котором он набрасывал нерв нервно-мышечного препарата на мышцу стеклянным крючком так, чтобы тот касался поврежденного и неповрежденного участков мышцы. В этом случае мышца тоже сокращалась. Таким образом, во втором опыте Л. Гальвани убедительно доказал существование «животного электричества», которое позже было зарегистрировано К. Маттеучи (1838) с помощью гальванометра. Одну из клемм прибора он присоединял к поврежденному участку мышцы, другую — к неповрежденному, при этом стрелка гальванометра отклонялась. Размыкание цепи прибора сопровождалось возвращением стрелки гальванометра в прежнее (нулевое) положение. В настоящее время используется внутриклеточная и внеклеточная регистрация электрических потенциалов с помощью монополярных, биполярных, мультиполярных электродов (металлических и стеклянных). В случае моно- полярного отведения один электрод активный, другой — индифферентный, площадь которого в десятки раз больше площади активного электрода. При внутриклеточном отведении применяется стеклянный микроэлектрод, представляющий собой микропипетку с кончиком диаметром 0,5—1 мкм (рис. 3.1, Б). Микроэлектрод заполнен ЗМ КС1, в его широкую часть вставлена тонкая про-
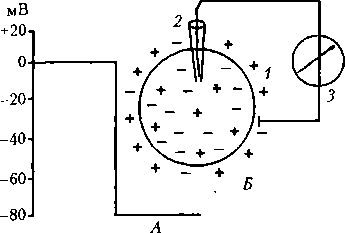
Рис. 3.1. Потенциал покоя возбудимой клетки (Л) и схема опыта его регистрации (Б):
1 — клетка; 2 — микроэлектрод; 3 — регистратор волока (например, серебряная), соединенная с регистрирующим устройством. В качестве индифферентного внеклеточного электрода часто используют хлорированную серебряную пластинку. Микроэлектродный способ регистрации биопотенциалов позволил изучить механизмы создания электрических зарядов клеткой, возникновения возбуждения в живых клетках. Правда, еще задолго до появления микроэлектродной техники (конец XIX в.) стало ясно, что «животное электричество» обусловлено процессами, происходящими на клеточной мембране (Герман, Дюбуа-Реймон, Бернштейн). В настоящее время довольно хорошо изучены механизмы формирования мембранного потенциала покоя (ПП) и ПД, т.е. процесса возбуждения клетки, которое в эксперименте можно вызвать с помощью электрического раздражения. Сущность процесса возбуждения заключается в следующем. Все клетки организма имеют электрический заряд, создаваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для разных ионов и работы ионных насосов. При действии раздражителя на возбудимую клетку сначала, как правило, обычно очень быстро повышается проницаемость ее мембраны для Na+ и быстро возвращается к норме, затем — для К+ и тоже быстро, но с некоторым отставанием возвращается к норме. Вследствие этого ионы перемещаются в клетку и из нее согласно электрохимическому градиенту (совокупность концентрационного и электрического градиентов) — это и есть процесс возбуждения. Возбуждение возможно только в том случае, если клетка постоянно поддерживает ПП (мембранный потенциал) — характерный признак живой клетки. 3.2. Потенциал покоя Общая характеристика. ПП— это разность электрических потенциалов между внутренней и наружной средой клетки в состоянии ее покоя. При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом скачком отклоняется вниз и показывает отрицательный заряд внутри (см. рис. 3.1, А). Величина ПП обычно варьирует в пределах —30—90 мВ (в волокнах скелетной мышцы —60—90 мВ, в нервных клетках —50—80 мВ, в гладких мышцах —30—70 мВ, в сердечной мышце —80—90 мВ). Органеллы клеток имеют собственные заряды, величина которых вариабельна. Например, у митохондрий этот потенциал составляет 175 мВ. Роль П П: является основой для возникновения возбуждения (ПД), с помощью которого нервная система воспринимает и перерабатывает информацию, регулирует деятельность внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения мышцы. Нарушение процессов возбуждения в кардиомиоцитах ведет к остановке сердца, к прекращению сокращений гладких мыШЦ> влияет на транспорт ионов, а с ними и воды; ПП — это признак жизни самой клетки и организма в целом. Непосредственной причиной существования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки (Ю. Бернштейн, А. Ходжкин, Э. Хаксли, Б. Катц, 1902—1952). Таблица 3.1 Внутри- и внеклеточная концентрация Внутриклеточная |
Внеклеточная | |||||||||||||
| Na+ | 12 | Na+ | 145 | ||||||||||||
| К+ | 155 | К+ | 4 | ||||||||||||
| сг | 4 | СГ | 80 | ||||||||||||
| нсо3- | 8 | НСО3- | 27 | ||||||||||||
| А-* | 155 | Прочие катионы | 5 | ||||||||||||
* А — высокомолекулярные внутриклеточные анионы.
В нервных и мышечных клетках концентрация К+ внутри клетки примерно в 40 раз больше, чем вне клетки; концентрация Na+ вне клетки в 14 раз больше, нежели в клетке (табл. 3.1). Ионов СГ вне клетки в 20 раз больше, чем внутри клетки. В клетке имеется небольшое количество ионов Mg2+. Кальций в свободном (ионизированном) состоянии находится в основном вне клетки. Он содержится также в эндоплазматическом ретикулуме — в гиалоплазме его очень мало. Это отчасти обусловливается транспортом Са2+ наружу через клеточную мембрану, отчасти — поглощением его эндоплазматическим ретикулумом (это резервуар для Са2+) и другими органеллами, например митохондриями, связыванием Са2+ цитратом, глутаматом. В клетке находятся также крупномолекулярные анионы, главным образом это глутамат, аспартат, а также органические сульфаты, фосфаты.
Неравномерное расположение ионов внутри и вне клетки является, во-первых, следствием неодинаковой проницаемости клеточной мембраны для различных ионов, и во-вторых, — работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки их электрохимическим градиентам.
Проницаемость клеточной мембраны — это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильтрации. Проницаемость определяется, во-первых, размерами пор клеточной мембраны или каналов (управляемых, с воротным механизмом, и неуправляемых, без воротного механизма, и размерами частиц; во-вторых— растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липидов и непроницаема для пептидов). Термин «проводимость» следует использовать только лишь применительно к заряженным частицам. Следовательно, проводимость — это способность заряженных частиц (ионов) проходить через клеточную мембрану согласно электрохимическому градиенту.
Известно, что ионы, как и незаряженные частицы, диффундируют через мембрану из области с высокой концентрацией в область с низкой концентрацией. При большом градиенте концентрации и хорошей проницаемости мембраны, разделяющей соответствующие растворы, проводимость ионов может быть высокой, при этом наблюдается их односторонний ток. Если разность концентраций ионов по обе стороны мембраны снизится, то проводимость ионов также уменьшится, хотя проницаемость мембраны сохранится прежней — высокой. Кроме того, проводимость иона при неизменной проницаемости мембраны зависит и от заряда иона: одинаково заряженные ионы отталкиваются, а с противоположными зарядами — притягиваются. Возможна ситуация, когда при хорошей проницаемости мембраны проводимость ионов через последнюю оказывается низкой или нулевой в случае отсутствия движущей силы — электрохимического градиента.
Следует подчеркнуть, что проводимость иона зависит от его электрохимического градиента и от проницаемости мембраны, чем они больше, тем лучше проводимость иона через мембрану. Перемещения ионов в клетку и из нее осуществляются согласно концентрационному и электрическому градиентам в состоянии покоя клетки, преимущественно через неуправляемые (без воротного механизма) каналы, которые называются также каналами утечки ионов. Неуправляемые каналы всегда открыты, они практически не меняют своей пропускной способности при электрическом воздействии на клеточную мембрану и ее возбуждении. Однако движение ионов в клетку и из нее зависит и от электрического заряда клетки: одинаково заряженные ионы отталкиваются, а с противоположными зарядами — притягиваются.
Роль проницаемости клеточной мембраны и различных ионов в формировании ПП. В состоянии покоя клетки К+ выходит из нее значительно больше, чем входит Na+, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости ее для Na+. Органические анионы из-за своих больших размеров вообще не могут выходить из клетки, поэтому внутри клетки, пребывающей в состоянии покоя, отрицательных ионов оказывается больше, чем положительных. По этой причине клетка внутри имеет отрицательный заряд. Основные анионы, находящиеся в клетке, — это глутамат, аспартат, органические фосфаты, сульфаты.
Важно отметить, что во всех точках клетки отрицательный заряд практически одинаков. Об этом свидетельствует одинаковая величина ПП при введении микроэлектрода на разную глубину внутрь клетки, что имело место в опытах Ходжкина, Хаксли и Катца. Гигантский аксон кальмара (диаметр около 1 мм) в этом опыте находился в морской воде; один электрод вводили в аксон, другой — помещали в морскую воду. Заряд внутри клетки является отрицательным как абсолютно (в гиалоплазме клетки содержится больше анионов, нежели катионов), так и относительно наружной поверхности клеточной мембраны. Однако превышение абсолютного числа анионов над числом катионов в клетке чрезвычайно мало. Но этого различия достаточно для создания разности электрических потенциалов внутри и вне клетки и для возникновения ПД при раздражении клетки (см. п. 4.3).
Калий является основным ионом, обеспечивающим формирование ПП. Об этом свидетельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксона кальмара солевыми растворами. При уменьшении концентрации К+ в перфузате ПП уменьшается, при увеличении концентрации К+ — увеличивается. В покоящейся клетке устанавливается динамическое равновесие между числом выходящих из клетки и входящих в нее ионов К+. Электрический и концентрационный градиенты противодействуют друг другу- согласно концентрационному градиенту, К+ стремится выйти из клетки, а отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновешиваются, число выходящих из клетки ионов К+ становится равным числу входящих в клетку ионов К+. В этом случае на клеточной мембране устанавливается так называемый равновесный К-потенциал. Равновесный потенциал для любого иона можно рассчитать по формуле Нернста. Внеклеточная концентрация положительно заряженного иона приводится в числителе, внутриклеточная концентрация этого иона — в знаменателе; для отрицательно заряженных ионов расположение этих параметров противоположное:
г М>
Еу =------ log "
х ZF [X].
где Ех— потенциал, создаваемый данным ионом (A); R — газовая постоянная (8,31 Дм); Т— абсолютная температура (273 + 37 °C); Z— валентность иона; Е— постоянная Фарадея (9,65 х 104); [А]о — концентрация иона во внешней среде клетки (outside); [А], — концентрация иона внутри клетки (inside).
Равновесный потенциал для К+ при температуре 37°C с учетом соотношения концентрации его снаружи и изнутри (1/39) и валентности 1 равен —97мВ. Однако реальный ПП миоцита теплокровного животного несколько больше — около —90 мВ. Это объясняется тем, что в создании ПП принимают участие и другие ионы, хотя их роль менее значительна в сравнении с ролью иона К+. Равновесный потенциал для Na+ равен +55 мВ. В целом ПП — это алгебраическая сумма равновесных потенциалов всех ионов, находящихся внутри и вне клетки, и поверхностных зарядов клеточной мембраны, которые имеют гликопротеиды, фосфолипиды и гликолипиды.



