Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Первичный резервуар патогенных для человека микроорганизмовСодержание книги
Поиск на нашем сайте
Предыстория проблемы. Простейшие и их паразиты. Границы феномена сапронозного существования патогенных для людей микроорганизмов. Предадаптация к макрофагам. Исторические свидетельства.
Для прорывов в науке важно уметь выявлять артефакты, т. е. отдельные природные явления, не укладывающиеся в общепринятые научные представления. Собственно задачей ученого и является выявление таких артефактов и затем их объяснение. Но по сложившейся в науке практике в этом случае он рискует нажить себе много неприятностей, и, прежде всего, обвинений в «ненаучности». Чем банальней «научность», тем меньше, к сожалению, она вызывает к себе критическое отношение ученых. Однако если артефакт существует в объективной реальности, он неизбежно обвалит господствующую концепцию при дальнейшем совершенствовании методологии исследований. Ниже мы очень кратко рассмотрим ряд таких артефактов, меняющих наши представления о первичных резервуарах возбудителей опасных болезней человека, важных уже с точки зрения планирования противоэпидемических мероприятий, но одновременно необходимых и для понимания роли иммунной системы позвоночных в распространении ретровирусов. Предыстория проблемы. Выдающийся немецкий гигиенист Макс Петтенкофер (Pettenkofer Max, 1818–1901) известен еще и тем, что решительно возражал против ведущей роли «заноса» микробного фактора в этиологии холеры как пандемической болезни. По Петтенкоферу, если холерный зародыш обозначить буквой X, а благоприятную для его развития почву буквой Y, а происходящий от их взаимодействия яд буквой Z, то ни X, ни Y не могут сами по себе вызывать холеру, а только один Z, т. е. яд. При этом специфическая природа яда определяется специфическим зародышем, а количество яда свойствами почвы. Благоприятной для развития яда, по мнению Петтенкофера, была почва, в верхних своих слоях пористая и проницаемая для воздуха и воды, и загрязненная в то же время отбросами органических веществ. Если холерный зародыш заносится в такую местность, где почва обладает данными свойствами, то он начинает созревать, обусловливая эпидемическое развитие болезни. Напротив, в тех местностях, где почва не обладает упомянутыми свойствами, занесение холерного зародыша не ведет к дальнейшему распространению болезни (Петтенкофер М., 1885). Направление в эпидемиологии, связывающее развитие эпидемических болезней со свойствами почвы, называлось тогда локализмом. Петтенкофер не был ни голословен, ни одинок в своих взглядах. Один из последователей Петтенкофера в России, профессор Казанского университета Н. К. Щепотьев (1884), исследуя географию появления вспышек чумы в Астраханской области, пришел к выводу, что для объяснения эпидемического распространения чумы «еще недостаточно одной переносчивости ее». По его наблюдениям, существуют местности, в которые чума не заносится никогда и ни при каких обстоятельствах. Для развития же эпидемии необходимо временное и местное появление еще особого фактора, независимого от чумного агента. Только с появлением этого фактора открывается возможность чумному агенту фиксироваться, развиваться и существовать в данной местности. С исчезновением этого фактора исчезает и чума; а чумный агент, выделенный больными организмами, быстро разрушается. Фактор должен иметь в различное время различную степень интенсивности и экстенсивности. Разность поражения чумой одной и той же местности в различные годы и в различные месяцы одного и того же года обусловливается именно различной степенью напряженности действия этого неизвестного фактора. Его природа определяется «совокупностью наблюдения над движением и развитием эпидемий». Щепотьев считал, что развитие чумного агента зависит от теплоты и влажности почвы. Какой-то еще не распознанный продукт разложения органических веществ почвы, образовавшийся под влиянием определенных физико-химических процессов, и составляет фактор X, столь необходимый для эпидемического развития чумы (Супотницкий М. В., Супотницкая Н. С., 2006). К числу сторонников Петтенкофера относился знаменитый патолог того времени Р. Вирхов (Virchow; 1821–1902). Но в конечном итоге его взгляды на доминирующую роль неизвестных науке факторов почвы (Y) в развитии эпидемий большая часть ученых проигнорировала. После открытия микроорганизмов — возбудителей инфекционных болезней — «вес» набирало другое направление, контагионистическое (Р. Кох, Г. Гафки и др.), видевшее только в контактной передаче микроорганизмов причину возникновения инфекционных болезней у людей. Бактерии прекрасно «состыковывались» со средневековым учением о контагии. Но теперь стало ясно, что это живой организм (contagium vivum), а не «яд», и что его можно получать в большом количестве и изучать в лабораторных условиях. У ученых появилась новая положительная мотивация — возможность разрабатывать вакцины, сыворотки, диагностические препараты и пр., и никто не обязан был верить теоретическим выкладкам ученого — реликта добактериологической эпохи. Сам Петтенкофер окончил жизнь самоубийством, а его фамилия в бактериологии стала нарицательной и упоминалась в ХХ столетии лишь в связи со случаем, когда он, чтобы доказать непричастность холерных вибрионов к холерным эпидемиям, выпил холерную культуру. В общем, был такой ретроград — Макс Петтенкофер, не верил в очевидное, в то, что холерные пандемии вызываются холерным вибрионом, тем и запомнился. Причина научного поражения Петтенкофера и других локалистов заключалась не в отсутствии у них аргументов своей правоты, с этим все обстояло скорее наоборот (см. «Исторические свидетельства»). Как правило, локалисты представляли своим оппонентам обширные и убедительные описания эпидемических процессов и примеры медицинской статистики. Читатель может найти некоторые из них в книге Ф. Ф. Эрисмана (1893) и убедиться в том, что сегодня на таком уровне эпидемиологический анализ уже не проводят. Дело тут было в используемой локалистами методологии — они не могли инструментально продемонстрировать факторы Х, Y и Z. Их взгляды на эпидемический процесс на фоне достижений бурно развивающейся тогда медицинской бактериологии стали выглядеть умозрительными, а обнаруженные особенности таких процессов, необъяснимые как передача «контагия», считаться артефактами. И вообще в эпидемиологии с открытием микроорганизмов — возбудителей инфекционных болезней — все стало как бы понятно и ясно. Поэтому локалистические представления подверглись не опровержению, а забвению, как уже ненужные. А микроорганизмы, возбудители инфекционных болезней людей и животных, отдельные авторы до конца 1930-х гг. продолжали называть контагиями. Для контагионистов различия в условиях существования микроорганизмов в естественных условиях и в питательном бульоне в лаборатории, носили лишь количественный характер (концентрация и соотношение питательных веществ, температура среды, содержание кислорода и т. п.). В рамках этого подхода для них не существовало методических ограничений еще почти 100 лет. Методический уровень бактериологии, необходимый для экспериментального обоснования экологических позиций локалистов и позволяющий изучать патогенные для людей микроорганизмы в водных и почвенных экосистемах, не был достигнут не только в конце ХIХ столетия, но и почти на всем протяжении двадцатого. Оставшись без оппонентов, ученые-контагионисты уже не стремились искать иные причины появления эпидемий и пандемий инфекционных болезней вне общих рассуждений о возможности «заноса» их возбудителей-контагиев. Эпидемиологи, сами того не подозревая, вернулись к взглядам средневековых врачей где-то времен после «черной смерти» (1346–1351). С конца XIX столетия в эпидемиологии и микробиологии господствуют антропоцентристские представления о причинах существования в природе патогенных микроорганизмов. Они очень просты и хорошо запоминаются студентами — все патогенные микроорганизмы поддерживаются в природе дикими животными и от них передаются людям, а затем распространяются между людьми. Когда реальная эпидемиология инфекционной болезни не вписывалась в эту схему, ее просто придумывали. «Выдающимися» примерами такого подхода стали объяснения холерных пандемий заносом больными холерного вибриона из холерных местностей и возрождение раннесредневековых взглядов на эпидемиологию чумы, как на болезнь, распространяемую кораблями. Правда, теперь роль переносчика «чумного контагия» играли не вещи больных чумой, а инфицированные крысы. Нельзя утверждать, что противоречий и «пробелов» в этих представлениях никто не замечал. Артефакты накапливались и требовали объяснения. Еще в 1956 г. W. Drozanski описал облигатные внутриклеточные паразиты свободно живущих амеб. Тогда эти микроорганизмы назвали Sarcobiumlyticum, но в последствии было установлено, что они относятся к опасному для людей семейству бактерий Legionella и их реклассифицитовали. Сегодня они известны как Legionellalytica. В 1958 г. В. И. Терских на основе своих наблюдений заново обосновал положение о том, что внешняя среда может служить средой обитания патогенных микроорганизмов. Под давлением эпидемиологических наблюдений Mollaret H. (1963), первым среди чумологов, был вынужден вернуться к забытому в начале XX столетия учению Макса Петтенкофера (правда, не упоминая его имени), предполагающему участие почвы в поддержании в природе возбудителей опасных инфекционных болезней. Смысл его гипотезы сводится к тому, что чумной микроб при наличии соответствующих условий может длительно персистировать в почве нор грызунов (теллурическая чума). Развивая гипотезу Mollaret, М. Балтазар (1964) пришел к заключению, что цикл чумы в природных очагах состоит из двух фаз: паразитической (на грызунах и их блохах — кратковременной и неустойчивой); и непаразитической (существование в почве нор — устойчивой). Однако где находится первичный резервуар возбудителя, эти исследования не прояснили. Чумологи по-прежнему рассуждали о заносе чумы кораблями и о тому подобных «научно обоснованных» фактах. Прошла незамеченной работа С. В Никульшина с соавт. (1993), показавших способность ряда амеб фагоцитировать Y.pestis и сохранять ее в предцистах. Механизм и эпидемическая значимость этого явления оставались непонятным до открытия явления «некультивируемости бактерий» и разработки методов молекулярной диагностики. Суть феномена «некультивируемости» заключается в следующем. Исследователи обнаруживают микроорганизмы в одноклеточных животных (простейших) методами молекулярной диагностики, но не могут подтвердить их наличие культивированием на искусственной питательной среде. С антропоцентристской точки зрения феномен объяснялся просто — случайностью; микроорганизм случайно попал в неблагоприятную для него среду (благоприятная среда, разумеется, питательный бульон, приготовленный в лаборатории этих исследователей) и находится в состоянии стресса. Однако границы феномена оказались значительно более широкими, чем это можно ожидать от «случайности». Простейшие и их паразиты. Свободноживущие амебы имеют, по крайней мере, две стадии развития: трофозоиты (trophozoite) — вегетативные метаболически активные формы; и цисты (cyst) — «спящие формы», позволяющие им выжить в неблагоприятных условиях среды. Отдельные амебы, такие как Naegleria spp., имеют дополнительную стадию флагеллят (flagellate stage), другие, такие как Mayorella и Amoeba, включают виды, не формирующие цисты (рис. 21).
Рис. 21. Стадии развития Hartmanella vermiformis
А. вегетативная форма; Б. циста. Черная полоса внизу фотографии соответствует 2 микронам (Greub G., Didier R., 2004).
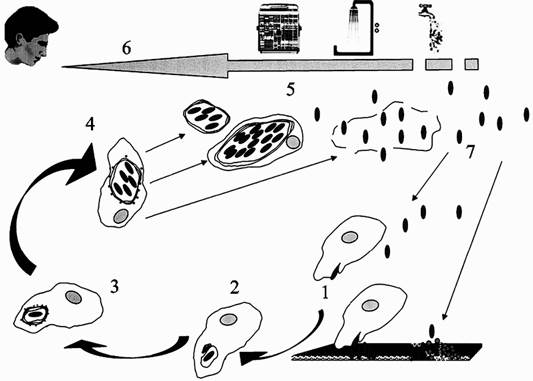
Свободноживущие амебы представлены повсеместно и могут быть изолированы из воздуха, почвы, воды, назального секрета позвоночных. В почве они наиболее распространены в тех ее участках, которые покрыты растениями. На растениях обильно паразитируют грибы и бактерии, являющиеся пищей для свободноживущих амеб (Rodriguez-Zaragoza S., 1994). Более подробно о таксономии, анатомии и экологии простейших можно прочитать в работах Г. Н. Калкинса (1912), Л. Н. Серавина (1984), К. Хаусмана (1988) и А. П. Пехова (1994). Нас же простейшие интересуют как эволюционные предки макрофагов и как первичный резервуар возбудителей опасных инфекций для человека. Амебы живут в широком интервале условий окружающей среды. Адаптация бактерий к протозойным хищникам происходит уже миллиарды лет, и в настоящее время ученые насчитывают не менее четырех форм их взаимодействия, хотя, естественно, эти знания являются только предварительными. Во-первых, отдельные бактерии могут использовать их для увеличения своей численности в окружающей среде, как это показано для Klebsiellaaerogenes, размножающихся «за счет» Acanthamoeba castellanii. Во-вторых, некоторые бактерии продуцируют литические компоненты, разрушающие простейших, и тем самым предотвращающие фагоцитоз, например Bacillusliqueniformis синтезирует литический компонент, направленный против Nagleria fowleri. В-третьих, между бактериями и простейшими могут устанавливаться эндосимбиотические отношения. Известно, что бактерии могут оставаться в простейших длительное время в некультивируемом состоянии. Было установлено экспериментально, что паразитические бактерии могут существовать в простейших в таком состоянии не менее шести лет. В-четвертых, в процессе эволюции и коэволюции бактерии могут совершенствовать механизм внутриклеточного существования в амебах и затем, после проникновения в организм позвоночных, пользоваться этим механизмом для выживания в макрофагах (например, Legionellapneumophila; см. ниже) (Harb O. et al., 2000). В основе взаимодействия простейших с микроорганизмами (бактерии, дрожжи, вирусы и др.) лежит способность группы их поверхностных рецепторов взаимодействовать со структурами, богатыми углеводом маннозой. Они получили общее название — маннозные рецепторы. Их присутствие на поверхности амебы необходимо для связывания микроорганизмов, инициирования их поглощения клеткой и доставки в лизосомы для переваривания (Allen P. G., Dawidowicz E. A., 1990). Позже было показано, что и сами одноклеточные паразиты проникают в эпителиальные клетки высших животных посредством взаимодействия с маннозными рецепторами этих клеток. Например, посредством такого механизма Acanthamoeba проникает в роговицу глаза человека и вызывает кератит (Zhantao Yang et al., 1997). Механизмы проникновения свободноживущих почвенных амеб в организм позвоночных также сложно опосредованы с рецепторными структурами внеклеточного матрикса. Например, почвенная амеба Balamuthiamandrillaris, возбудитель смертельного для человека гранулематозного амебного энцефалита (granulomatous amoebic еncephalitis), использует для проникновения в мозг три типа компонентов внеклеточного матрикса: коллаген I, главный компонент соединительной ткани; молекулы фибронектина, имеющие критическое значение для адгезионных процессов; и ламинин-1 (laminin-1), ключевая молекула для формирования базального слоя. Распознание ламинина зависит от галактозо-связывающего белка (Rocha-Azevedo B. et al., 2007). Типичные молекулы данного типа в эукариотических клетках — галектины (galectins) относятся к семейству галактозных лектинов и широко распространены среди различных клеток животных (более подробно о галектинах см. в работе Leffler H. et al., 2004). Весьма любопытно в аспекте понимания эволюции иммунной системы то обстоятельство, что амебы в условиях in vitro реагируют на фактор некроза опухолей (TNF), интерлейкин-1бета (IL-1бета), интерлейкин-8 (IL-8) и циклооксигеназу-2 точно также, как нейтрофилы и макрофаги — т. е. как на хемоаттрактанты (Blazquez S. et al., 2006). С филогенетической точки зрения, направленное движение клеток в ответ на внешние раздражители, является давно известной биологической реакцией (New D. C., Wong J. T., 1999). Следовательно, сигнальные молекулы, которые мы сегодня, путаясь в определениях, называем хемокинами и цитокинами, а также их рецепторы стали «средством общения» между простейшими задолго до появления нейтрофилов и макрофагов как клеток иммунной системы позвоночных организмов. Уже в этом десятилетии обнаружены микроорганизмы — внутриклеточные паразиты простейших, не растущие на искусственных питательных средах, но являющиеся патогенными для человека. Например, T. J. Marrie et al. (2001) установили, что такие микроорганизмы, как LLAPs (Legionella-like amoebal pathogens — возбудители пневмоний), Parachlamydia acanthamoeba BN9 (болезнь Кавасаки) и Afipia felis (болезнь кошачьей царапины), паразитирующие в свободноживущих почвенных амебах, очень хорошо размножаются в человеческих моноцитах, но не растут на искусственных питательных средах. Если "пойти в обратную сторону", т. е. проверить способность уже охарактеризованных на искусственных питательных средах патогенных для человека микроорганизмов паразитировать внутри макрофагов, непрофессиональных фагоцитов и даже в нефагоцитирующих клетках (фибробластах, эпителиоцитах), то можно обнаружить, что все они способны к такому паразитизму. Подготовленный О. В. Бухариным (1999) ретроспективный обзор таких экспериментов включает не менее 20 патогенных для человека видов бактерий. Более поздний обзор G. Greub, D. Raoult (2004) включает уже около 50 видов бактерий и отдельные виды риккетсий и вирусов. Экспериментальные данные также свидетельствуют о другом феномене паразитизма патогенных для человека микроорганизмов у простейших — упрощении генома паразита при специализации его к своему одноклеточному хозяину. Например, сравнение геномов Yersinia pseudotuberculosis и Y. pestis для ученого, привыкшего считать, что патогенность микроорганизмов обусловлена приобретением генов факторов патогенности (т. е. усложнением генома), требует преодоления некоторого психологического барьера. Возбудитель чумы, более патогенный для человека и большинства модельных животных, чем возбудитель псевдотуберкулеза, утрачивает значительную часть генов, которые традиционно относят к генам вирулентности и патогенности. По данным, обобщенным А. П. Анисимовым (2002), по сравнению с псевдотуберкулезным микробом, Y. pestis утрачивает гены адгезинов, уреазы (сдвиг рамки считывания), инвазинов Inv и Ail (вставка IS-элементов), подвижности, способности к синтезу О-боковых цепей ЛПС (не установленные механизмы образования мутаций) и ряд других. Из 17 «биосинтетических» генов, выявленных у псевдотуберкулезного микроба, 5 в геноме Y. pestis инактивированы за счет вставок и делеций. Компьютерный анализ полного генома чумного микроба (штамм CO92, биовар Orientalis; выделен от человека, погибшего от легочной чумы) показал наличие 149 псевдогенов. Объяснение этому феномену я вижу в сравнении экологии обоих микробов. У псевдотуберкулезного микроба очень широкий круг хозяев, в основном среди гидробионтов. Он является комменсалом для зоопланктона (дафнии, циклопы), бентосных животных (кольчатые черви, моллюски, личинки насекомых и др.) и высших растений. Возбудитель же чумы специализирован на узком круге почвенных простейших. Следовательно, упрощение его генома является следствием дегенеративной эволюции, характерной для видовой специализации любого паразита. Но специализации не к отдельным видам позвоночных животных (включая человека), а к простейшим. Границы феномена сапронозного существования патогенных для людей микроорганизмов. О том, что границы этого феномена очень широки, свидетельствует много косвенных факторов. Например, все так называемые возбудители опасных и особоопасных инфекционных болезней (чумы, мелиоидоза, сапа, туляремии, сибирской язвы, бруцеллеза, лихорадки Ку и др.), одновременно являющиеся и потенциальными агентами биологического оружия, имеют много сходства в эпидемиологии, биологических свойствах и в клинике вызываемых ими поражений людей, объяснить которое возможно только в том случае, если предположить, что они являются истинными паразитами (или эндосимбионтами) почвенных простейших. В пользу такого предположения говорит «привязанность» вызываемых ими вспышек болезней к конкретным местностям; отсутствие у них резистентности детоксикационного типа к антибиотикам (ненужной для внутриклеточного паразитизма); вовлечение в инфекционный процесс лимфатических узлов и фагоцитирующих клеток крови; а также «запутанность» вопроса о факторах их патогенности и токсинах. Ни один из обнаруженных у них таких «факторов» не может однозначно в эксперименте прояснить клинику вызываемой микроорганизмом у людей болезни, больше похожую на реакцию организма на суперантигенный раздражитель. Не обладают они и контагиозностью, т. е. способностью передаваться от одного теплокровного организма к другому при контакте, что обычно предполагает воздушно-капельный механизм передачи возбудителя болезни непосредственно от одного заболевшего к другому. Редкие случаи «перехода» неконтагиозной бубонной чумы в контагиозную вторично-легочную лишь исключения, подтверждающие вышеприведенные наблюдения. Ни больные с такой формой чумы, ни появившиеся через инфицирование от них больные с первично-легочной чумой, не способны поддерживать возбудитель чумы в природе, так как в 100 % известных случаев (без лечения антибиотиками) они погибают. И если этим возбудителям инфекционных болезней не придумывать эпидемиологию (см. «Исторические свидетельства»), то понять, как они существуют в природе без резервуара среди простейших, практически невозможно. Vibriocholerae. Возбудитель холерыявляется членом семейства Vibrionaceae, о котором известно, что оно прекрасно размножается в простейших A. polyphaga и N. gruberi. Их выживание в пределах цист N. gruberi предполагает то, что свободноживущие амебы могут длительно сохранять холерный вибрион при неблагоприятных условиях среды (Thom S. et al.,1992). Достоверно установлены биоценотические связи холерного вибриона, обитающего в воде, с водными организмами, в том числе и с растениями. Например, была показана способность ряски, зоопланктона (Islam M. et al., 1990) и низших ракообразных (Голубев Б. П.,1993) поддерживать высокую концентрацию холерного вибриона. Islam et al. (1990) также выявили активное размножение и длительную (до 15 месяцев!) персистенцию некультивируемых вибрионов Эль Тор в культуре сине-зеленых водорослей, которые признаются ими возможным резервуаром холеры в межэпидемические периоды. Марамович А. С. и Наркевич М. И. (1993) пришли к выводу о том, что водный гиацинт является природным резервуаром холеры Эль Тор. Он обеспечивает существование вибрионов в межэпидемические периоды и способствует реализации водного пути передачи возбудителя. Бухарин О. В., Литвин В. Ю. (1997) экспериментально показали, что водный гиацинт способствует размножению холерных вибрионов. Их концентрация в его стеблях и листьях была в 300 раз выше, чем в воде. Эти и другие данные позволили Пушкаревой В. И. и Литвину В. Ю. (Пушкарева В. И., Литвин В. Ю.,1994; Пушкарева В. И.,1994), выдвинуть гипотезу о клонально-селекционном механизме изменения бактерий в почвенных и водных сообществах, способном обеспечивать формирование эпидемически значимых вариантов возбудителей сапронозов в их природных резервуарах. Гипотеза весьма интересная, поэтому ее стоит привести подробно. В процессе естественной циркуляции холерных вибрионов среди гидробионтов — хозяев возбудителя в водоемах, имеет место селекция токсигенных клонов и их накопление в бактериальной популяции при благоприятных условиях (активном пассировании через хозяев). Всего же у холерных вибрионов в водной экосистеме формируется, как минимум, две экологические ниши. Одна из них (непосредственно водная среда) наиболее характерна для авирулентных штаммов. Вторую нишу (сообщество водных организмов) населяет преимущественно вирулентная часть микробной популяции, устойчивая к перевариванию в организме гидробионтов — первичная функция токсигенности холерных вибрионов, возможно, как раз и состоит в защите бактериальной популяции, обитающей в водоемах, от хищничества простейших и других гидробионтов. Их объем относительно друг друга меняется. Например, при изменении численности хозяев или сдвигах в структуре водного сообщества. В разных условиях и в разные сезоны в водной популяции вибрионов могут доминировать то токсигенные, то атоксигенные клоны холерных вибрионов. Изменение уровня токсигенности всей популяции вибрионов во времени происходит за счет выхода в водную среду нарастающего числа токсигенных вибрионов из организма погибших инфузорий, где они накапливаются благодаря селективному преимуществу перед атоксигенными и слаботоксигенными вибрионами. Прогревание воды до температуры 20 0С и резкое увеличение трофности водоемов в июле — августе определяют пик численности холерных вибрионов, тесно сцепленный с пиком численности планктона. Интенсивно пассируясь среди гидробионтов, вибрионы окончательно выходят из покоящегося состояния (число бактериологически высеваемых культур максимально); численность и вирулентность водной популяции вибрионов резко возрастает, достигая эпидемически значимых показателей. Именно к этому периоду неизменно приурочен пик заболеваемости людей в очагах умеренных широт. Первичные и независимые случаи инфицирования холерой людей, связаны с водоемами, после чего распространение холеры может принять и вторичный (эстафетный) характер, в виде классической вспышки (по работе Бухарина О. В. и Литвина В. Ю., 1997). Приведенная выше клонально-селекционная теория состыковывается с гениальными прозрениями Макса Петтенкофера (1885) следующим образом. Если принять нетоксигенные клоны холеного вибриона за фактор Х, то гидробионты, через которые пассируются токсигенные клоны возбудителя холеры, можно считать тем фактором Y, существование которого Петтенкофер так настойчиво отстаивал, а сами токсигенные клоны представляют собой фактор Z. Однако только ли для холерного вибриона верна эта теория? Легионеллы. Для легионелл возможность сапронозного существования установлена бактериологическими методами еще в 1950-х гг. Вспышки легионеллезов обычно сопровождаются высокой смертностью среди заболевших людей. Поэтому экология легионелл и механизмы их проникновения в человеческие популяции находятся в поле зрения ученых уже не менее 30 лет, с момента печально известной вспышки болезни в Филадельфии в 1976 г. В данной работе легионеллы рассматриваются как «опережающий объект» в исследованиях сапронозов, что предполагает перенесение выявленных при их изучении закономерностей на менее исследованные микроорганизмы, обитающие в почвенных и водных экосистемах. В пределах водных сообществ легионеллы существуют в ассоциации с планктоном или как составная часть биопленок (Rogers J. et al., 1994). Разные легионеллы инфицируют своих протозойных хозяев по разным механизмам, что говорит за многообразие экологических отношений в мире микроорганизмов и простейших. Амебы имеют критическое значение для существования легионелл в окружающей среде и для их способности вызывать инфекционную болезнь у людей (Harb O. et al., 2000). Схематическое изображение взаимодействия L. pneumophila и одноклеточных животных в окружающей среде и механизма ее трансмиссии к человеку приведено на рис. 22.
Рис. 22. Механизм взаимодействия L. pneumophila и одноклеточных животных в окружающей среде и их трансмиссии к человеку
1. L. pneumophila из биопленок, формируемых совместно с другими бактериями или из их суспензий, инфицирует Рrotozoa. Проникновение в простейшие осуществляется по механизму фагоцитоза.2. После проникновения в амебу L. pneumophila остается в мембранно-связанной вакуоле, которая рекрутируется органеллами, такими как митохондрии, и не сливается с лизосомами.3. В пределах 4 ч после проникновения, вакуоль с L. pneumophila окружается эндоплазматическим ретикулумом.4. L. pneumophila реплицируется в пределах специализированной вакуоли.5. Точный механизм «выхода» бактерий из простейших неизвестен, но считается, что возможна их экскреция в составе везикул. Могут лизироваться и сами амебы.6. Передача L. рneumophila к людям осуществляется механически, через воздух, выбрасываемый кондиционерами, с каплями воды в душевых и т. п. способами.7. Legionellae могут длительное время выживать в окружающей среде. Они повторно инфицируют Protozoa или реколонизируют биопленки (Harb O. et al., 2000).
Взаимоотношения легионелл и Protozoa, нашедшие свое отражение в патогенности этих бактерий для людей, могут быть следующими. Амебы после превращения в цисты, создают бактериям надежное убежище от враждебных условий внешней среды (температура, влажность, дезинфектанты и т. п.). Этим можно объяснить способность легионелл распространяться и населять инженерные системы зданий, прошедшие специальную обработку. К тому же, реплицирующаяся в амебах L. рneumophila становится почти в 1000 раз более резистентной к антимикробным соединениям, чем при культивировании на искусственных питательных средах (Barker J., Brown M. R., 1995). Жизненный цикл L. pneumophila вамебах сходен с таковым в макрофагах (см. «Предадаптация к макрофагам»)(рис. 23).
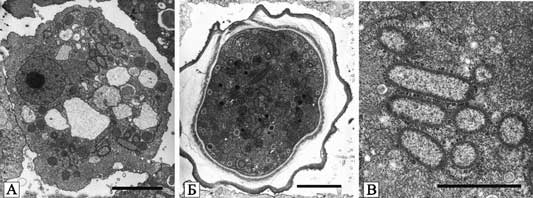
Рис. 23. Фагоцитоз L. pneumophila амебой H. vermiformis в условиях in vitro
А. адгезия; Б. поглощение; В. интернализированные бактерии. mi — митохондрия. Черная маркерная полоса соответствует 0,5 мк (Greub G., Didier R., 2004).
Пассирование легионелл в амебах повышает их инвазивность и вирулентность для человека. В пользу последнего предположения говорят экспериментальные данные, показавшие многократно возросшую способность L. рneumophila, выращенных в простейших, внедряться в макрофаги мышей и реплицироваться в легких по сравнению с теми же легионеллами, выращенными на искусственной питательной среде (Cirillo J. D. et al., 1994). Отдельные гены, необходимые для выживания L. рneumophila в макрофагах, такие как dot/icm, pmi и mip, также нужны им для выживания в амебах (табл. 7).
Таблица 7. Гены L. pneumophila, необходимые для инфицирования и внутриклеточного существования в Protozoa и макрофагах [10]
Гены | Функция | Роль в инфекции | Примечания dot/icm (defect in organelle trafficking и intracellular multiplication) | Аппарат секреции IV типа. Конъюгационный перенос ДНК. | Правильное созревание фагосомы. Репликация в Protozoa. Цитотоксичность, вызванная формированием пор | Agrobacterium tumefaciens и Bordetellapertussis, используют подобную систему секреции для переноса ДНК (virB) и секреции коклюшного токсина (ptl), соответственно pmi (protozoan and macrophage infectivity) | Неизвестная функция | Внутриклеточное выживание. Правильное созревание фагосомы. Присоединение к клетке-хозяину | Вместе с dot/icmобеспечивают L. рneumophila возможность выживать и реплицироваться в эволюционно значительно отстоящих клетках pilBCD (encodes a prepillin peptidase) | Биогенез пилей IV типа | Секреция II типа | Секреция бактериальных белков, необходимых для внутриклеточного выживания | Аппарат секреции II типа позволяет L. рneumophila выживать в Protozoa и в клетках млекопитающих rpos | Стационарная фаза ответа на стресс | Регуляция генов, необходимых для внутриклеточного выживания | — prp | Гомологичен prpD-гену S. typhimurium | Внутриклеточное выживание | — mip (macrophage infectivity potentiator) | Относится к семейству белков пептидил-пролил изомераз (peptidyl-prolyl cis/trans isomerases, — PPIase) | Внутриклеточное выживание в течение ранних стадий инфекции | Необходим для инфицирования макрофагов, эпителиальных клеток и простейших emi (early macrophage-induced locus) | Неизвестная функция | То же | — asd (aspartate-b-semialdehyde dehydrogenase) | Биосинтез диаминопимелата | Внутриклеточная репликация | — Гены усвоения железа | Захват и ассимиляция внутриклеточного железа | То же | -
Способность легионелл реплицироваться в амебах показывает то, что амебы являются для этих бактерий самой адекватной питательной средой. Chlamydophilapneumoniae. Бактерия хорошо известна как причина различных респираторных болезней и атеросклероза. Сероэпидемиологические исследования обнаружили ассоциацию антител к С. pneumoniae с атеросклерозом коронарных, сонных и церебральных артерий, а также инфарктом миокарда (Lee Ann Campbell et al., 1998). В природе бактерия поддерживается в Acanthamoebacastelanii, но не в Parachlamydiaceae или Simkaniaceae (Essig A. et al., 1997). Микобактерии. Для Mycobacterium leprae — микобактерии, вызывающей болезнь, уже тысячи лет известную под названием проказа, было показано, что она может поддерживаться в свободноживущих почвенных амебах. Однако бактериальной репликации или микробного лизиса амеб, вызванного этими микобактериями, описано не было, что свидетельствует в пользу их симбиотических отношений и частично объясняет клинику болезни. Развитие проказы у людей происходит в течение десятков лет, при этом инкубационный период болезни может длиться до пяти лет. У микобактерий, вызывающих туберкулез, отношения с простейшими могут складываться как по типу эндосимбиоза, так и как паразитические. В экспериментах, выполненных в условиях in vitro, установлена способность M. avium, M. marinum, M. ulcerans, M. simiae и M. habane проникать в свободно живущие почвенные простейшие. M. smegmatis, M. fortuitum и M. рhlei размножаются в самых различных амебах и вызывают их лизис. Хорошо изучены взаимодействия между M. avium и Acanthamoeba. Было установлено, что M. аvium, выращенная в амебах, более вирулентна чем такая же микобактерия, выращенная на искусственной питательной среде. Причины этого явления следующие. Во-первых, M. аvium, выращенная в амебах, более активно инфицирует как сами амебы, так и клеточные линии интерстинального эпителия (HT-29) и макрофаги. Во-вторых, такие M. аvium обладают расширенными возможностями по колонизации кишечника мышей, в минимальных дозах вызывают у них микобактериальную инфекцию, и быстро проникают в печень и селезенку. M. avium при неблагоприятных условиях среды хорошо выживает в цистах Acanthamoeba. В то же время M. avium, живущая в амебах, более резистентна, чем M. avium, находящаяся в макрофагах, к антимикробным препаратам, используемым для профилактикимикобактериоза у больных СПИДом (рифабутин, кларитромицин и азитромицин) (Greub G., Didier R., 2004). Микобактерии, вызывающие у людей туберкулез, могут существовать длительное время как эндосимбионты простейших. Например, Zhatao Yang et al. (2007) обнаружили в контактных линзах амебы вида Acanthamoebalugdunensis. Ими было установлено, что эти амебы уже не менее 6 лет поддерживают существование микобактерий (предположительно, M. avium или M. Intracellulare). Причем сами микобактерии не оказывали на Acanthamoeba никакого цитопатического эффекта (рис. 24).
Рис. 24. Электронная микрофотография Acanthamoeba KA/LC6 с бактериями-эндосимбионтами
A. Бактерии-эндосимбионты палочкообразной формы, случайным образом распределенные в цитоплазме трофозоита; Б. Цисты с эндосимбионтами. В. Увеличенные бактерии-эндосимбионты в цитоплазме трофозоитов. По поверхности бактерий-эндосимбионтов «рассыпаны» рибосомы. Черная маркерная полоса соответствует 2 мк. По Zhatao Yang et al. (2007).
Механизм проникновения патогенных микобактерий в популяции людей значительно более сложен, чем у легионелл. В цепочке передачи возбудителя болезни появляется новый промежуточный хозяин — личинки кровососущих комаров. Инфицированные личинки через воду попадают в организм животных и птиц, а от них к человеку. Полученные данные согласуются с сообщениями о выделении патогенных и атипичных микобактерий из проб воды, взятых из прудов, канав и луж, и используемой для поения крупно рогатого скота, а также из опилок, соломы и сена (Ермакова М. с соавт.,1995). Burkholderiaceae. Бактерия B.cepacia ассоциирована у людей с несколькими легочными инфекциями, особенно с цистическим фиброзом. Роль свободноживущих почвенных амеб как резервуара B. сepacia пока не доказана, однако их участие в трансмисси возбудителя возможно, так как они способны выбрасывать в окружающую среду везикулы, заполненные B. cepacia (Marolda C. L. et al., 1999). B. pseudomallei —возбудитель опасной болезни, мелиоидоза, и потенциальный агент биологического оружия. Показан
|
||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 113; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.11 (0.021 с.) |