
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цветоразличение на уровне сетчатки
Первая физиологическая информация о различении цветов на клеточном уровне была получена спустя 250 лет после открытия И. Ньютона в исследованиях Г. Светихина (1956), который на костистой рыбе осуществил внутриклеточную регистрацию активности нейронов сетчатки — сначала он принял их за колбочки, но они оказались горизонтальными клетками. На освещение сетчатки эти клетки отвечали только медленными потенциалами (потенциалов действия не наблюдалось). Светихин обнаружил три типа клеток: первый тип, названный им L-клетками, гиперполяризовался при световой стимуляции независимо от спектрального состава света, второй тип, названный r-g-клетками (красно-зелеными), гиперполяризовался волнами малой длины с максимумом ответа на зеленый свет и деполяризовался волнами большой длины с максимумом ответа на красный свет; третий тип, названный с учетом теории Геринга г/-Ь-клетками (желто-синими), отвечал по типу клеток r - g, но с максимумом гиперполяриза-ции на синий и максимумом деполяризации на желтый свет. У клеток r - g и у-Ь белый свет вызывал лишь слабые и быстро затухающие ответы, как и следовало ожидать ввиду широкополосного спектрально-энергетического состава белого света. Кроме того, у клеток этих типов, которые можно назвать оппонентно-цветовыми клетками, свет с некоторой промежуточной длиной волны не вызывал никакой реакции. Поскольку эти клетки реагируют на окрашенный, но не на белый свет, они, вероятно, связаны с цветовыми ощущениями. 264 Глава IV Психофизиология зрительного восприятия 7 Неиробиологня цветового зрения 265
Р. Де Валуа и его сотрудники (1958) зарегистрировали ответы клеток наружного коленчатого тела макаки, поразительно сходные с ответами элементов сетчатки костистой рыбы на цветовые стимулы. Ранее с помощью поведенческих тестов было показано, что цветовое зрение макаки и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и, по-видимому, вполне правомерно сравнение психофизиологических механизмов восприятия цвета у человека и макаки. Р. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки скрещения, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки скрещения до другого конца спектра. И вновь аналогия с цветовыми процессами Геринга была полной: Де Валуа выявил оппонентно-цветовые клетки двух типов, красно-зеленые и желто-синие; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки скрещения, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты опытов на обезьянах особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки скрещения точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».
Следующий этап состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки. У рецептивных полей большинства оппонентно-цветовых клеток Де Валуа обнаружилась удивительная организация. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию. Центр мог быть типа on - или off -. В типичном случае центр представлен исключительно красными, а тормозящая периферия исключительно зелеными колбочками. Поэтому при красном как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порожда- ют сильное торможение с о//-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа. Реакция на диффузный свет — в данном случае on-реакцйи на красный, о//-реакции на синий и зеленый и отсутствие реакции на белый свет — показывает, что такая клетка должна отвечать на цветовой стимул. Но реакция на цветовые края и отсутствие ответов на диффузный свет говорит о том, что эта клетка связана и с восприятием формы. Эти клетки получили название «тип 1».
В четырех верхних слоях были найдены клетки еще двух типов. Клетки «типа 2» составляют около 10% нейронной популяции и имеют рецептивные поля, состоящие только из центра. Повсюду в этом центре у некоторых клеток выявляется красно-зеленая, а у остальных — сине-желтая оппонентность. Рецептивные поля еще примерно у 15% клеток в четырех верхних слоях коленчатого тела и у всех клеток в двух нижних крупноклеточных слоях имеют центр и периферию, но эти клетки не проявляют цветовых предпочтений; создается впечатление, что центр и периферия их полей получают одинаковый относительный вклад от всех трех видов колбочек. Эти клетки считаются широкополосными и называются клетками «типа 3». Все эти данные согласуются с моделью Геринга: кроме классов клеток с цветовой оппонентностью есть также третий класс, не обладающий этим свойством, но с широкополосной пространственной оппонентностью. Вентральная (нижняя) пара слоев коленчатого тела отличается от четырех дорсальных слоев тем, что состоит исключительно из клеток с широкополосными центрами полей. Эти клетки проявляют любопытную форму цветовой оппонентности. Центры их полей в несколько раз крупнее, чем центры нейронов мелкоклеточных слоев, и у них есть ряд других интересных особенностей. Предполагается, что эти клетки обслуживают отделы мозга, играющие важную роль в восприятии формы, глубины и движения. Большинство типов клеток, описанных для нейронов коленчатого тела, встречается также и в сетчатке. В коленчатом теле они больше обособлены, что облегчает их изучение. 7.3.3. Цветочувствительные нейроны коры Эксперименты, которые выполнялись на нейронах зрительной коры, показали, что лишь немногие клетки чувствительны к цвету. При картировании рецептивных полей при помощи черных и белых полосок переход к цветовым стимулам давал точно такой же резуль- 266 Глава IV Психофизиология зрительного восприятия 7 Неиробиология цветового зрения 267
В 1978 г. появились данные, полученные в результате гистохимических исследований зрительной коры: при окрашивании коры для выявления фермента цитохромоксидазы — фермента, который есть во всех клетках и участвует в клеточном метаболизме — в верхних слоях появлялась неоднородность — темноокрашенные участки шириной около 250 мк чередовались с неокрашенными шириной около полумиллиметра. На препаратах срезов первичной зрительной коры, выполненных параллельно ее поверхности, выявился узор, напоминающий горошек. Эти неоднородности получили название «пузырьки». В нейрофизиологических экспериментах важно было произвести регистрацию электрической активности нейронов, образующих эти «пузырьки», исследовать структуру их рецептивных полей и цветочувствительность. Результат оказался совершенно неожиданным. Проходя расстояние в 0,25 мм, равное диаметру пузырька, можно исследовать примерно пять или шесть клеток. Всякий раз при пересечении пузырька попадавшиеся на пути электрода клетки были полностью лишены ориентационной избирательности, что заметно контрастировало с высокой ориентационной избирательностью клеток, расположенных вне пузырьков. Этому отсутствию ориентационной специфичности можно было бы дать объяснение: эти клетки могли неизбирательно получать входные сигналы от соседних ориентационных клеток, лежащих вне пузырьков, и поэтому еще способны были отвечать специфической реакцией на линии (полоски и т.п.), но после объединения всех возможных ориентации всякое предпочтение какой-либо из них исчезало. Или могли быть похожими на клетки коленчатого тела или на корковые клетки слоя 4С и, таким образом, быть проще внепузырько-вых ориентационно-избирательных клеток. Вскоре был получен ответ на этот вопрос: оказалось, что большинство этих клеток имеет рецептивные поля с центром и периферией. Специально выполненные эксперименты показали, что многие из них участвуют в кодировании цвета. Больше половины пузырьковых клеток имели оппонентно-цве-
товые рецептивные поля с центром и периферией, но вели себя явно более сложным образом, чем клетки «типа 1» наружного коленчатого тела. Они практически не отвечали на белые пятна любых размеров и формы. Но на небольшие цветные стимулы, вспыхивающие в центре рецептивного поля, они энергично реагировали в одном диапазоне длин волны и затормаживались в другом диапазоне; некоторые активировались длинными волнами (красным светом) и подавлялись короткими (зеленым и синим светом), другие вели себя противоположным образом. Как и среди клеток коленчатого тела, можно было в зависимости от положения максимальных реакций на спектральной шкале выделить два класса клеток — красно-зеленые и сине-желтые (здесь «красный», «зеленый» и «синий» указывают на соответствующие типы колбочек, а слово «желтый» — на параллельные входы от красных и зеленых колбочек). Таким образом, эти клетки весьма напоминали оппонентно-цветовые клетки коленчатого тела, обладающие только центром («тип 2»). Но в отличие от клеток «типа 2» эти цветокодирующие пузырьковые клетки чаще всего отвечали на большие белые или цветные пятна, каков бы ни был спектральный состав света. Они вели себя так, как если бы центральная система рецептивного поля была у них окружена кольцом оппонентности. Если говорить о наиболее распространенном типе клеток, то центр типа r + g ~ (красный-оя, зеленый-q//), казалось, окружен у них кольцом типа rg ^ (красный-q//, зеленый-огс).
Эти клетки получили название двойные оппонентные из-за их красно-зеленой или желто-синей оппонентности в центре и антагонизма периферии по отношению к любой реакции в центре, будь она on - или off -. Поэтому они не отвечают не только на белый свет в любой геометрической конфигурации, но и на большие пятна, независимо от их спектрального состава. Центры рецептивных полей, как и у клеток «типа 2», были в несколько раз больше, чем у клеток коленчатого тела «типа 1». Вперемешку с двойными оппонентными клетками обоих классов (красно-зелеными и желто-синими) встречались и обычные широкополосные клетки с центром и периферией. Эти широкополосные клетки отличались от клеток верхних слоев коленчатого тела и от клеток слоя АС коры более крупными размерами своих центров. В настоящее время мы можем лишь гадать, какова схема связей двойных оппонент -ных клеток. Ясно, что каким-то образом двойная оппонентность должна достигаться: либо ее источник находится на нижних уровнях, либо это происходит в коре. Поэтому кажется, что исходное предположение о раздельном анализе цвета и формы как разных аспектов восприятия находит подтверждение в виде физического обособления пузырько- Fcbi 268 Глава IV Психофизиология зрительного восприятия 8 Два глаза 269
Открытия, сделанные в области механизмов восприятия цвета при объединении психофизических и нейрофизиологических методов, помогают понять такие явления, как результаты смешения цветов или константность цвета. 8. Два глаза Многие органы тела парные, однако глаза (как и уши) отличаются тем, что работают в тесном взаимодействии: они вместе воспринимают и сличают информацию, так что совместно выполняют работу, которая недоступна для одного глаза или уха. Воспринимаемые изображения размещаются в глазах на изогнутой поверхности сетчатки, однако, несмотря на это, их можно назвать двумерными. Удивительным в работе зрительной системы является ее способность синтезировать два различных изображения в единое целостное изображение объектов, расположенных в трехмерном пространстве. У некоторых животных зрительные оси направлены в противоположные стороны — это характерно для животных с латеральным расположением глаз. Переход в расположении глаз от бокового к фронтальному, благодаря чему стала возможной точная оценка расстояния, сыграл важную роль в тот период, когда у млекопитающих развивались передние конечности, способные держать предметы, манипулировать ими и цепляться за ветки деревьев. Для животных, которые прыгают с ветки на ветку, быстрая и точная оценка расстояния близких объектов очень важна, и работа обоих глаз, которые совместно дают стереоскопическое зрение, в высшей степени продуктивна. Кошка имеет фронтальное расположение глаз, работающих совместно, однако у них плотность фоторецепторов приблизительно одинакова по всей сетчатке.
На сетчатке животных с фронтальным расположением глаз сформировалось особое место, где плотность рецепторов наиболее высока — это фовеа. Она возникает только тогда, когда становится необходимой точная оценка глубины воспринимаемого изображения, как это имеет место у птиц или живущих на деревьях обезьян; у них развита фовеальная область сетчатки и существует точный контроль движений глаз. Стереоскопическое зрительное восприятие движений также обеспечивается парными фасеточными глазами насекомых Оно высоко развито у таких насекомых, как стрекоза, которая хватает свою добычу на лету на большой скорости. Фасеточные глаза неподвижно за- креплены на голове, и механизм их стереоскопического зрения проще, Стереоскопическое зрение (стереопсис) — это способность воспринимать глубину пространства и оценивать удаленность предметов от глаз. Наше знакомство с основами бинокулярного зрения начнется с выяснения вопроса о физиологическом соответствии обеих сетчаток. Если мы будем смотреть на любую звезду двумя глазами, а затем закроем один глаз, не меняя при этом положения головы, то кажущиеся направления, в каких нам до того виделись на небе звезды, не изменяются. Это означает, что при параллельно направленных зрительных линиях обоих глаз изображения удаленных предметов, падающих на сетчатку, видятся в одних и тех же местах пространства независимо от того, падают эти изображения на сетчатку правого глаза или на сетчатку левого глаза, или же на обе сетчатки сразу. Следовательно, некоторым местам одной сетчатки соответствуют определенные места другой сетчатки, т.е. раздражающие их объекты видятся нами в одних и тех же местах пространства. Такие места сетчаток носят название корреспондирующих точек. Они характеризуются тем, что возбуждение их дает ощущение одного объекта в поле зрения. Мы хорошо знаем, что все, что мы фиксируем, т.е. то, что направляем в центральную ямку сетчатки, видится нами одиночно. Поэтому прежде всего центральные ямки сетчатки следует признать корреспондирующими точками. Корреспондирующими точками сетчаток являются также и все те места сетчаток, которые лежат в одном и том же направлении и на одном и том же расстоянии от центральной ямки сетчатки. При каждом положении глаз этим корреспондирующим точкам сетчаток соответствуют строго определенные точки во внешнем пространстве. Совокупность всех точек пространства, которые дают изображение на корреспондирующих местах сетчаток, называется гороптером. Для разных положений глаз гороптер имеет разную форму. Раздражение корреспондирующих точек вызывает впечатление одиночного предмета. Что же происходит, если изображение предмета падает в обоих глазах на некорреспондирующие, диспаратные точки сетчатки? В этих случаях могут возникать двойственные изображения. В существовании их легко убедиться на следующем опыте Возьмем два тонких карандаша или лучше две спицы а и b и будем держагь перед глазами так, чтобы они находились на одной прямой, в медиальной плоскости головы и отстояли дру1 oi друга приблизительно на 15-20 см Будем фиксировать дальнюю спицу а в таком случае изображение от ближней спицы b упадет па диспаратные точки сетчатки (в чевом глазу - влево, в нравом глазу - вправо or центральной ямки) Сообразно с этим мы 270 Глава IV. Психофизиология зрительного восприятия 8. Два глаза 271
Но всегда ли в случае раздражения диспаратных точек возникает двоение? Оказывается, что не всегда. Если несоответствие раздраженных мест сетчатки не чрезмерно и если это несоответствие к тому же носит односторонний характер (т.е. в обоих глазах раздражение попадает только на левые или только на правые половины сетчатки), то вместо двоения у нас возникает новое впечатление — впечатление большей или меньшей удаленности данного объекта по сравнению с фиксируемым. Таким образом, известное несоответствие изображении на сетчатке дает нам возможность судить об относительной удаленности предмета. Существует взаимосвязь между двумя механизмами, описанными выше: конвергенцией глаз, которая служит своего рода дальномером, и различием между двумя изображениями, называемым диспаратнос-тью. Угол конвергенции является регулятором системы диспаратнос-ти. Когда глаза фокусируют отдаленный предмет, любая диспарат-ность между изображениями означает большие различия по глубине, чем в тех случаях, когда глаза конвергируют для восприятия близко расположенных объектов. Если бы этого не было, отдаленные предметы казались бы ближе друг к другу по глубине, чем близкие предметы, расположенные на том же расстоянии друг от друга, потому что дис-паратность тем больше, чем ближе находятся предметы. Действие механизма координации, компенсирующего эти геометрические соотношения, довольно легко наблюдать, если нарушить конвергенцию, сохранив прежнюю диспаратность. Мозговые механизмы, обеспечивающие стереоскопическое восприятие глубины, могут интегрировать наборы линий, воспринимаемые каждым глазом отдельно, синтезировать объекты из двух случайных структур и эффективно находить диспаратность. Бинокулярная конвергенция Глаза — парный орган. Очевидный интерес представляет вопрос: получает ли определенная корковая клетка входные сигналы от обоих глаз, и если да, то одинаковы ли эти входы в количественном и качественном отношении? Для того чтобы получить ответ, необходимо вернуться к наружному коленчатому телу (НКТ) и выяснить, есть ли на этом уровне ней- роны, имеющие входы от обоих глаз. НКТ — это самый низкий уровень, на котором уже возможно объединение сигналов от двух глаз на одной клетке. Однако возможность эта, по-видимому, здесь не реализована — два пучка входных волокон распределяются по разным кле-. точным слоям, между которыми нет или почти нет взаимодействия. Как можно было бы ожидать, учитывая раздельность проекций от двух глаз, отдельная клетка НКТ отвечает на стимуляцию одного какого-то глаза и совсем не реагирует на стимуляцию другого. Судя по результатам некоторых экспериментов, стимулы, подаваемые на «чужой» для данной клетки НКТ глаз, могут оказывать слабое воздействие на реакцию, вызываемую со «своего» глаза. Однако можно считать, что каждая клетка находится под контролем только одного глаза. Пути, идущие от каждого глаза, должны сойтись вместе, так как мы, глядя на что-либо, видим одну целостную картину. Повседневный опыт говорит о том, что если закрыть глаз, то большого изменения не произойдет — предметы будут казаться такими же четкими, такими же реальными и такими же яркими. Общее поле зрения для двух глаз будет несколько шире, поскольку каждый глаз видит на своей стороне большую область пространства, чем другой глаз, хотя эта разница составляет всего лишь около 20—30°. Существенное отличие бинокулярного зрения от монокулярного состоит в ощущении глубины. В коре обезьяны те клетки, которые получают входные сигналы от НКТ и имеют концентрические рецептивные поля, сходны с клетками НКТ в том, что они тоже монокулярные. На этом корковом уровне обнаружено примерно одинаковое число клеток, возбуждающихся от левого и от правого глаза, по крайней мере в тех участках коры, которые обслуживают область поля зрения в радиусе около 20° от центральной точки фиксации. Однако на следующем уровне коры выявляются уже бинокулярные клетки, простые и сложные, причем у обезьян больше половины таких клеток могут реагировать на сигналы от каждого из глаз независимо. При обнаружении бинокулярной клетки, можно очень точно сравнить ее рецептивные поля в обеих сетчатках. Сначала при закрытом правом глазе животного картируется рецептивное поле клетки в левом глазу. При этом определяется его точное положение на экране или на сетчатке, а также его сложность, ориентация и расположение возбуждающих и тормозных зон. При картировании рецептивного поля определяется его микроструктура — выясняется, простое оно или сложное, исследуется возможность реагировать на конец линии и дирекциональная чувствительность. После этого повторяется вся процедура измерений для правого глаза при закрытом левом. Оказалось, что у большинства бинокулярных клеток все свойства, выявляемые в 272 I лава IV Пспхофизиочогия зрительно! о восприятия 8 Два глаза 273
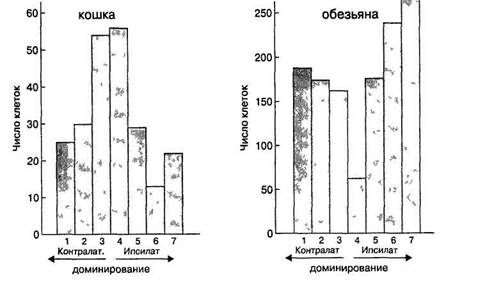
опытах с левым глазом, обнаруживаются и при стимуляции правого глаза — то же положение на сетчатке, та же дирекциональная чувствительность. Это позволяет заключить, что все связи, идущие к данной клетке от левого глаза, совпадают по структуре со связями, идущими от правого глаза Говоря о таком дублировании связей, нужно сделать одно уточнение Если, определив для клетки оптимальный стимул, его положение, ориентацию, направление движения и т д, сравнить ее ответы при стимуляции одного глаза и при стимуляции другого, интенсивность реакции не всегда окажется одинаковой Некоторые клетки действительно одинаково хорошо активируются от обоих глаз, однако другие явно дают более сильный разряд при стимуляции определенного глаза. В целом, за исключением той части корковых клеток, которые обслуживают периферию поля зрения, не обнаруживается никакого особого преимущества того или другого глаза — в каждом полушарии число клеток, лучше активируемых с противоположной стороны (от контралатерального глаза) и с той же стороны (от ипсилатерального глаза), примерно одинаково При этом встречаются все степени относительного доминирования глаза, начиная от клеток, возбуждаемых исключительно с левого глаза, и кончая клетками, отвечающими только на стимуляцию правого глаза Можно оценить численность различных групп нейронов, сформированных по принципу наибольшей эффективности Все изученные клетки можно произвольно разбить на классы, в зависимости от того, отвечают они на стимуляцию сразу двух глаз, только одного какого-либо глаза или же того и другого, но отдавая преимущество какому-то из них Получается семь классов по относительной эффективности воздействия на них того или другого глаза (рис 18) Результаты представлены на гистограммах для кошки и обезьяны. Здесь сразу налицо сходство и различие в распределении клеток у этих животных Видно, что у обоих видов бинокулярные клетки встречаются достаточно часто, причем среди клеток с односторонним доминированием хорошо представлены оба глаза, что у кошек бинокулярных клеток очень много, что у обезьян численность монокулярных и бинокулярных клеток примерно одинакова, причем у бинокулярных клеток достаточно сильно выражено доминирование одного глаза (группы 2 и 5) Гистограммы показывают, что реже всего встречаются клетки, испытывающие одинаковые влияния от каждого глаза Отвечают ли бинокулярные клетки при стимуляции обоих глаз лучше, чем при стимуляции одного глаза? Оказывается, многие клетки работают именно так — при стимуляции одного глаза они активируются слабо или не возбуждаются совсем, а при раздражении, посту- пающем от двух глаз, дают сильный импульсный разряд Особенно ярко это проявляется в том случае, когда оба глаза стимулируются одновременно и совершенно одинаково Эффект синергии у многих клеток выражен слабо или не отмечается вовсе — такие клетки отвечают на стимуляцию обоих глаз примерно так же, как и на стимуляцию каждого глаза по отдельности Рис 18 Распределение нейронов по группам глазодоминирования В первую группу входят клетки, на которые влияет только один контралатеральный глаз, во вторую — клетки, испытывающие влияние двух глаз, но преимущественно контралатерального, в третью и четвертую — отвечающие только при стимуляции сразу двух глаз и т д
8.2. Механизмы взаимодействия двух глаз Связи одиночных клеток с двумя глазами еще раз указывают на высокую степень специфичности соединений в мозгу. Входные системы связей данной клетки позволяют ей отвечать только на линию определенной ориентации и лишь на одно направление движения Кроме того, эти системы представлены двумя копиями, по одной от каждого глаза Как показывают опыты, выполненные на животных разного возраста, большинство связей, по-видимому, должно быть сформировано и готово к работе уже к моменту рождения животного 18-1015 274 Глава IV Психофизиология зрительного иосприятия 8 Два!лаза 275
Организация нейронов стриарной коры, проявляющих свойство глазодоминирования, наиболее интенсивно и продуктивно разрабатывалась в исследованиях Д. Хьюбела, Т. Визела, С.Ле Вея и других Опыты этих исследователей показали, что группы клеток стриарной коры с различной глазодоминантностью довольно многочисленные и были обнаружены раньше всех других функциональных организаций, объединявших нейроны, специализированные на выделении каких-либо других признаков. Разработано много методов исследования таких группировок, поэтому они сейчас лучше всего изучены. В опытах Д. Хьюбела и Т. Визела (1962) было обнаружено, что, когда микроэлектрод входит в кору перпендикулярно ее поверхности, он встречает одну за другой клетки, лучше реагирующие на стимуляцию одного и того же глаза. Если микроэлектрод ввести в другом месте в нескольких миллиметрах от предыдущего, снова для всех встречающихся клеток доминирующим будет один глаз — тот же, что и раньше, или другой В слое АС, имеющем входы непосредственно от НКТ, доминирование одного глаза становится уже не относительным, а абсолютным. В тех же слоях, которые расположены выше и ниже и находятся дальше в цепи синаптических переключений, более половины всех клеток могут возбуждаться и от недоминирующего глаза. Такие клетки называются бинокулярными. Если электрод вводили с наклоном к поверхности коры, как можно ближе к линии, параллельной поверхности, то клетки с разной глазодоминантностью чередовались — доминирующим был то один глаз, то другой. Полный цикл этой смены соответствовал примерно одному миллиметру. Очевидно, если бы можно было видеть структуру коры сверху, она представлялась бы мозаикой, составленной из чередующихся зон с доминированием правого и левого глаза. Чем обусловлено такое чередование, стало ясно после разработки нового метода окраски, позволяющего прослеживать ветвление отдельных аксонов, приходящих из НКТ, и распределение их окончаний в коре. Ветвление одного аксона таково, что тысячи его концевых синапсов образуют в слое АС два или три скопления шириной 0,5 мм, разделенных промежутками около 0,5 мм. Поскольку клетки НКТ монокулярные, каждый отдельный аксон связан в конечном счете либо с правым, либо с левым глазом Предположим, что аксон — это одно из волокон, приходящих от левого глаза; в этом случае каждое входящее в тот же участок коры волокно, связанное с левым г пазом, будет разветвляться в тех же скоплениях окончаний величиной ОД мм каждое Промежутки между этими скоплениями размером тоже 0,5 мм будут заняты концевыми разветвлениями волокон от правого глаза Такой особый тип распределения аксонов из НКТ в слое АС сразу позволяет объяснить строгую монокулярность клеток в этом слое. Для того, чтобы избирательно окрашивать только одно волокно, требовался новый метод, и он был предложен в конце 70-х гг. Этот метод основан на использовании аксонного транспорта — процесса, в результате которого различные материалы (белки или даже более крупные частицы) непрерывно переносятся в обоих направлениях внутри аксона. Некоторые из них транспортируются со скоростью нескольких сантиметров в час, другие — нескольких миллиметров в сутки. Для того, чтобы окрасить одиночный аксон, с помощью микропипетки в него вводят вещества, о которых известно, что они переносятся по аксону, но не влияют на структуру клетки (например, фермент пероксидазу из хрена) Фермент распространяется по аксону в обоих направлениях и катализирует химическую реакцию, продукт которой очень эффективно окрашивает клетку. Поскольку фермент служит катализатором, достаточно ничтожных его количеств, чтобы получить интенсивную окраску. Важно и то, что в самой нервной ткани фермента с подобными свойствами нет: это исключает возможность нежелательной окраски фона. Организацию нейронов в колонки глазодоминантности, проходящие в коре от поверхности мозга вплоть до белого вещества, подтвердили и морфологические данные: группы клеток в слое АС служат главными местами передачи зрительной информации в клеточные слои, лежащие выше и ниже этого слоя. Наличие некоторого числа горизонтальных и диагональных связей длиной около миллиметра, идущих по всем направлениям, должно приводить к некоторому размыванию четких зон доминирования правого или левого глаза в слоях, расположенных выше и ниже слоя АС Поэтому можно ожидать, что клетка, расположенная прямо над центром зоны доминирования левого глаза в слое 4, будет явно лучше отвечать на стимуляцию этого глаза, а, возможно, и полностью контролироваться им, тогда как клетка, расположенная ближе к границе между участками разной глазодоминантности, может оказаться бинокулярной без какого-либо доминирования одного из глаз. Действительно, при горизонтальном продвижении электрода в верхнем слое коры или же в слое 5 или 6 отмечаются постепенные изменения глазодоминантности' сначала будут встречаться клетки, намного сильнее активируемые одним глазом, затем клетки, у которых эта асимметрия выражена слабее, потом 18* 276 Глаиа IV Психофизиология зрительного восприятия 8 Диа глаза 277
|
||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-01-09; просмотров: 66; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.196.223 (0.061 с.) |
||||||||||||||||||||||||||||||||||||||||||




 7.3.2. Цветочувствительные нейроны НКТ
7.3.2. Цветочувствительные нейроны НКТ
 тат — не было влияния на интенсивность ответа цветовых стимулов. Однако у некоторых клеток, составлявших примерно десятую часть всех нейронов верхнего слоя коры, обнаруживались цветовые предпочтения. Клетки более отчетливо отвечали на предъявление цветных полосок, чем на белый и черный стимул. Тем не менее, большая часть клеток коры к цвету оказалась совершенно нечувствительной. Это удивительно, так как на более низком уровне — наружном коленчатом теле — очень большая часть клеток кодирует информацию о цвете и передает ее кортикальным нейронам. Непонятно, что же происходит с этой информацией при ее передаче на более высокий уровень обработки зрительных изображений.
тат — не было влияния на интенсивность ответа цветовых стимулов. Однако у некоторых клеток, составлявших примерно десятую часть всех нейронов верхнего слоя коры, обнаруживались цветовые предпочтения. Клетки более отчетливо отвечали на предъявление цветных полосок, чем на белый и черный стимул. Тем не менее, большая часть клеток коры к цвету оказалась совершенно нечувствительной. Это удивительно, так как на более низком уровне — наружном коленчатом теле — очень большая часть клеток кодирует информацию о цвете и передает ее кортикальным нейронам. Непонятно, что же происходит с этой информацией при ее передаче на более высокий уровень обработки зрительных изображений.




 вых и непузырьковых участков первичной зрительной коры. В областях выше стриарной коры — в зрительной зоне 2 и даже еще выше — эта обособленность сохраняется.
вых и непузырьковых участков первичной зрительной коры. В областях выше стриарной коры — в зрительной зоне 2 и даже еще выше — эта обособленность сохраняется.


 увидим спицу b в двойном виде. При этом двойственные изображения носят перекрестный характер. Левое изображение видится правым глазом, а правое — левым. Если же фиксировать ближнюю спицу Ь, то раздвоится дальняя спица а. Возникающие в этих условиях двойственные изображения будут одноименными. Правый глаз будет видеть правое изображение, а левый — левое. Подобное различие в характере двоения объясняется тем, что раздражения, падающие на сетчатке влево от центральной ямки, мы относим к предметам, находящимся вправо от фиксируемой точки.
увидим спицу b в двойном виде. При этом двойственные изображения носят перекрестный характер. Левое изображение видится правым глазом, а правое — левым. Если же фиксировать ближнюю спицу Ь, то раздвоится дальняя спица а. Возникающие в этих условиях двойственные изображения будут одноименными. Правый глаз будет видеть правое изображение, а левый — левое. Подобное различие в характере двоения объясняется тем, что раздражения, падающие на сетчатке влево от центральной ямки, мы относим к предметам, находящимся вправо от фиксируемой точки.




 (по Хьюбелу)
(по Хьюбелу)





 8.2.1. Колонки глазодоминантности
8.2.1. Колонки глазодоминантности клетки с равной зависимостью от обоих глаз, после чего начнет усиливаться влияние другого глаза. Такие плавные переходы контрастируют с той внезапной сменой глазодоминантности, которая наблюдается при движении электрода. Если смотреть «сбоку», две совокупности клеток в слое АС выглядят как чередующиеся участки. Однако исследователям хотелось узнать, как будет выглядеть распределение этих участков при взгляде на поверхность коры сверху. Если бы они были отмечены на поверхности коры черным или белым цветом, то их пространственное размещение могло быть представлено несколькими разными способами: в шахматном порядке, в виде чередующихся черных и белых полос, в виде черных пятен на белом фоне или же могли быть использованы любые комбинации этих паттернов.
клетки с равной зависимостью от обоих глаз, после чего начнет усиливаться влияние другого глаза. Такие плавные переходы контрастируют с той внезапной сменой глазодоминантности, которая наблюдается при движении электрода. Если смотреть «сбоку», две совокупности клеток в слое АС выглядят как чередующиеся участки. Однако исследователям хотелось узнать, как будет выглядеть распределение этих участков при взгляде на поверхность коры сверху. Если бы они были отмечены на поверхности коры черным или белым цветом, то их пространственное размещение могло быть представлено несколькими разными способами: в шахматном порядке, в виде чередующихся черных и белых полос, в виде черных пятен на белом фоне или же могли быть использованы любые комбинации этих паттернов.


