
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биполярные и горизонтальные клетки
Горизонтальные и биполярные клетки вместе с амакриновыми клетками расположены в среднем слое сетчатки. Биполярные клетки занимают в сетчатке стратегическую позицию, поскольку все сигналы, возникающие в рецепторе и поступающие к ганглиозным клеткам, должны пройти через них. Это означает, что они входят в состав как прямых, так и непрямых путей. В отличие от этого горизонтальные клетки входят только в непрямые пути. Горизонтальные клетки встречаются намного реже биполярных, которые в целом преобладают в среднем слое. Биполярная клетка посылает по направлению к рецепторам единственный дендрит. Он либо образует синапс с одним рецептором (всегда с колбочкой), либо расщепляется на веточки, синаптически контактирующие с несколькими рецепторами. Если с одним биполя-ром связаны два или более рецепторов, они совместно занимают сравнительно малый участок сетчатки. Возникает вопрос: каковы синапсы между рецепторами и биполярными клетками — возбуждающие они, тормозные или обоих типов? Биполярные клетки, подобно рецепторам и горизонтальным клеткам, не генерируют импульсов, но здесь будут рассматриваться ои-от-веты (реакция активации на включение стимула), подразумевая деполяризацию в ответ на световой стимул и соответственно усиление вы- деления медиатора в выходных синапсах, и off -ответы (реакция активации на выключение стимула), подразумевая гиперполяризацию и уменьшение выброса медиатора. Что касается входных синапсов, передающих биполярам сигналы от рецепторов, то у биполяров с off -ответами они должны быть возбуждающими, поскольку сами рецепторы выключаются (гиперполяризуются) светом; у биполяров с ои-отве-тами входные синапсы должны быть тормозными. Чтобы понять, почему это так, следует лишь представить себе влияние малого светового пятна. Рецепторы активны в темноте, а свет вызывает гиперполяризацию и уменьшает их активность. Если синапс возбуждающий, то рецептор активируется в темноте, а инактивируется на свету; если же синапс тормозный, биполяр в темноте тормозится, а свет, выключая рецептор, снимает это торможение, т.е. биполярная клетка активируется (это действительно нелегко сразу понять). Является ли рецепторно-биполярный синапс возбуждающим или тормозным, зависит либо от выделяемого рецептором медиатора, либо от типа каналов в постсинаптической мембране биполярной клетки. В настоящее время есть много данных в пользу того, что биполяры двух типов имеют различные рецепторы.
Горизонтальные клетки важны потому, что они составляют ту часть непрямого пути, о которой больше всего известно. Это крупные клетки, и в нервной системе они принадлежат к числу самых странных. Их отростки тесно контактируют с окончаниями многих фоторецепторов, распределенных по площади, размеры которой велики по сравнению с участком, непосредственно связанным с одиночной биполярной клеткой. Каждый рецептор контактирует с обоими типами клеток второго порядка — биполярными и горизонтальными. Имеется несколько подтипов горизонтальных клеток, которые могут сильно различаться у разных животных; их самое необычное свойство, общее с амакриновыми клетками, — это отсутствие типичного аксона. Каким образом нейрон без аксона будет передавать информацию другим нейронам? Когда начали использовать в нейроанатомии электронный микроскоп, то выяснили, что дендриты могут образовывать синапсы на других нейронах, обычно на их дендритах. И наоборот, иногда окончания аксонов могут быть постсинаптическими структурами по отношению к другим аксонам, которые на них оканчиваются. Отростки, отходящие от тел горизонтальных и амакриновых клеток, могут выполнять функции как аксонов, так и дендритов. Своеобразны и синапсы, образуемые горизонтальными клетками с рецепторами: у них отсутствуют электронно-микроскопические признаки, обычно указывающие, в каком направлении передаются сигналы. Есть высокая вероятность того, что это синапсы с бидирекцио- 220 I лава IV Психофизиология зрительного восприятия 3. Сетчатка 221
То, что рецепторы передают горизонтальным клеткам информацию с относительно обширного участка сетчатки, означает, что рецептивные поля этих клеток должны быть большими. Опыты это подтвердили: величина их полей примерно соответствует размерам рецептивных полей биполярных или ганглиозных клеток, включая центр вместе с периферией. Они однородны, и освещение их в любом месте вызывает гиперполяризацию, которая тем сильнее, чем ярче световое пятно. Многие данные указывают на то, что горизонтальные клетки ответственны за периферию рецептивных полей биполярных клеток; кандидатов на эту роль, в сущности, нет, поскольку это единственные клетки, связанные с рецепторами на столь обширном пространстве. Там, где горизонтальные клетки непосредственно контактируют с биполярами, синапсы, передающие информацию ои-биполя-рам, должны быть возбуждающими (ввиду тормозящего влияния на периферию), а к о//-биполярам — тормозными. Если же влияние осуществляется через рецепторы, синапсы должны быть тормозными. 3.4. Амакриновые клетки Эти клетки удивительно разнообразны по форме и используют необычно большое число нейромедиаторов (идентифицировано более двадцати). Амакриновые клетки имеют ряд особенностей. Их тела расположены в среднем слое сетчатки, а отростки — в синаптической зоне, которая следует за этим слоем. Они образуют связи с биполярными и с ганглиозными клетками и таким образом создают между ними альтернативный, непрямой путь. И, наконец, у них нет аксонов, но зато их- дендриты способны к образованию пресинаптических окончаний на других клетках. Амакриновые клетки выполняют много различных функций, большей частью неизвестных. Один их тип, по-видимому, участвует в спе- цифических реакциях на движущиеся объекты (эти клетки обнаружены в сетчатке лягушки и кролика); другой тип участвует в создании пути, связывающего ганглиозные клетки с теми биполярами, которые имеют входы от палочек. Нет данных об участии амакриновых клеток в организации центра и периферии рецептивных полей ганглиозных клеток, но нельзя полностью исключить такую возможность. Функции большинства их форм остаются невыясненными: можно сказать, что анатомические сведения об амакриновых клетках значительно богаче физиологических. 3.5. Биполярные и ганглиозные клетки В 1976 г. удалось зарегистрировать реакции ганглиозных клеток кошки. Они были идентифицированы как клетки с on - или off -ответа- ми, затем через микроэлектрод ввели вещество, окрашивающее все дендритное дерево. Сравнив разветвления дендритов у клеток двух типов, исследователи увидели четкое различие: две совокупности дендритов оканчивались на двух отдельных субслоях в пределах синаптической зоны между средним слоем и слоем ганглиозных клеток. Дендриты клеток с о//-ответом всегда оканчивались ближе к среднему слою сетчатки, а дендриты клеток с ои-ответом — дальше от него. К тому времени уже было доказано, что биполярные клетки двух классов, образующие с рецепторами синапсы разной формы, отличаются также и расположением своих аксонных окончаний: у одних аксоны оканчиваются там, где кончаются дендриты ганглиозных клеток с ои-центром, а у других — там, где кончаются дендриты клеток с off -ответом. Таким образом, стало возможным реконструировать весь путь от рецепторов к ганглиозным клеткам для обеих систем — с on - и с о//-ответами. В результате был установлен неожиданный факт: на прямом пути именно система с о//-ответами имеет возуждающие синапсы на каждом уровне — от рецепторов к биполярам и от биполяров к ганглиозным клеткам, тогда как в системе с on-ответами синапсы между рецепторами и биполярами тормозные.
Разделение биполярных и гаглиозных клеток на категории с on - и о//"-ответами наверняка должно иметь свои корреляты в восприятии. Клетки с о//-ответом отвечают на темное пятно точно таким же образом, как клетки с оп-центром — на светлое пятно. Темнота так же реальна, как свет, и теперь мы находим биологическую основу этой реальности. Во многих сенсорных системах используются оппонентные пары: тепло — холод, черное — белое, поворот головы налево — направо, желтое — синее и красное — зеленое. Причина этого, вероятно, в характере нервной импульсации и ее изменений. В принципе можно
222 Глава IV Психофизиология зрительно! о восприятия '3 Сеччатка 223
3.6. Тормозное взаимодействие в элементах сетчатки Существование столь разветвленных и сложных синаптических связей на уровне элементов сетчатки дает возможность осуществлять тонкие процессы, которые лежат в основе обработки сигналов, уже в начале пути зрительной информации к высшим отделам мозга.
Взаимодействие возбуждающих и тормозных влияний, осуществляющееся с помощью связей между элементами сетчатки, приводит к таким особенностям распределения активности в зрительном нерве, которые не являются простыми и непосредственными копиями внешнего образа. Из огромного количества деталей временного и пространственного распределения освещения на мозаике рецепторов сетчатки отбирается определенная важная информация. Эта информация усиливается за счет менее важной информации и лишь после этого передается в центральную нервную систему. К числу наиболее важных особенностей распределения освещения относятся места перехода от одной интенсивности к другой и от одного цвета к другому. Действительно, даже если представлены только контуры, например, на штриховом, или контурном рисунке, значительная часть важной информации оказывается сохраненной. Исследователями описан механизм нервной интеграции, участвующий в обнаружении и усилении таких контуров. На глазе Limulus рассмотрено тормозное взаимодействие среди элементов сетчатки. Limulus — это краб, своеобразное живое ископаемое, который живет на восточном побережье США. Рецепторы его глаза непосредственно связаны с отдельными нервными волокнами. Нервные волокна этого древнего краба оказались наиболее пригодными для проведения исследований. Боковой глаз Limulus представляет собой сложный глаз, содержащий около 1000 омматидиев1. Нервные волокна отходят от омматидиев в
виде небольших пучков и образуют зрительный нерв. Сплетение нервных волокон связывает эти пучки сразу же за слоем омматидиев Каждый омматидий содержит около дюжины клеток: пучок клиновидных ретинулярных клеток и один биполярный нейрон — эксцентрическую клетку. Как эта клетка, так и ретинулярные клетки имеют аксоны, которые вместе составляют небольшой пучок, отходящий от оммати-дия; оба типа аксонов сильно ветвятся, образуя сплетение латеральных связей. Из зрительного нерва можно выделить небольшой пучок, содержащий одно активное волокно. Этот пучок помещают на электроды для регистрации нервных импульсов. Х.К. Хартлайн и его коллеги доказали, что регистрируемые на таком препарате импульсы возникают в эксцентрической клетке, которая, по-видимому, представляет собой нейрон, а не истинный рецептор. Эта клетка посылает дистальный дендритный отросток к центру рабдома, состоящего приблизительно из дюжины тесно соприкасающихся друг с другом рабдо-меров, которые представляют собой специализированные части ретинулярных клеток и, как полагают, являются фоторецепторами. Весь этот комплекс клеток, по-видимому, функционирует как одна «рецеп-торная единица». Было обнаружено, что в рецепторных клетках глаза Limulus частота импульсов связана приблизительно логарифмической зависимостью с интенсивностью света. Активность любого волокна в ответ на раздражение омматидия, от которого оно отходит, имеет особенности, типичные для реакций многих сенсорных нервов: между раздражением и появлением первого импульса протекает довольно длительный латентный период; интенсивность реакции зависит главным образом от интенсивности раздражения. Реакция на резкое повышение или понижение уровня раздражения протекает несколько сложнее.
Освещение других омматидиев, расположенных поблизости от омматидия, активность которого регистрируется, не вызывает разряда импульсов в аксоне его эксцентрической клетки. В любом одиночном волокне зрительного нерва можно вызвать активность, освещая лишь ту специфичную рецепторную единицу, от которой отходит это волокно. Тем не менее, сенсорные элементы глаза Limulus оказывают существенное влияние друг на друга через сплетение латеральных связей. Это влияние чисто тормозное. Частота разряда импульсов в волокне зрительного нерва, отходящем от данного омматидия, понижается, а иногда эти разряды совсем исчезают при освещении соседних областей глаза. Суть тормозного эффекта можно представить себе следующим образом. Способность омматидия давать разряд импульсов, идущих по аксону соответствующей эксцентрической к четки, уменьшается при 224 Гпава IV Психофизиолсиия зриюиьного восприятия 3 Сачагка 225
Рецепторы глаза Limulus оказывают друг на друга взаимные тормозные влияния. Активность каждого омматидия влияет на активность соседних омматидиев и в свою очередь зависит от их состояния. При регистрации активности двух волокон зрительного нерва, отходящих от двух не слишком удаленных друг от друга омматидиев глаза, частота разрядов импульсов в ответ на стабильное освещение в этих волокнах ниже, когда оба омматидия освещаются одновременно, чем когда каждый из них освещается отдельно. Степень торможения каждого омматидия зависит лишь от степени активности другого. Таким образом, активность каждого омматидия является результатом суммации возбуждения, вызванного действующим на этот омма-тидий световым раздражителем, и торможения, оказываемого другим омматидием. Активность рецепторной единицы измеряют частотой разряда импульсов в соответствующем аксоне. Эта реакция определяется возбуждением, вызванным действием внешнего раздражителя на рецептор, и уменьшается любыми тормозными влияниями, которые могут действовать на рецептор вследствие активности соседних рецепторов. Активность системы взаимодействующих элементов можно описать достаточно точно без подробного указания на распределение освещенности на рецепторной мозаике, так как зависимость тормозных влияний подразумевалась в величинах порогов и тормозных коэффициентов Тем не менее ясно, что выраженная зависимость тормозных порогов и коэффициентов от расстояния между элементами вводит топографический фактор, который должен иметь большое значение в деятельности сетчатки Следовательно, любое подробное описание пространственных характеристик тормозного взаимодействия должно давать полную картину связей между этими тормозными параметрами и соответствующими расстояниями на рецепторной мозаике Наибольших эффектов контрастности можно ожидать на границе (или поблизости от границы) между слабо и ярко освещенными об-яастями сетчатки. Рецепторная единица, расположенная в слабо освещенной области поблизости от этой границы, будет тормозиться не только слабо освещенными, но и ярко освещенными соседними единицами Суммарное торможение такой единицы будет поэтому большим, чем суммарное торможение других слабо освещенных единиц, расположенных дальше от границы, и частота разряда этой единицы будет ниже, чем у последних единиц. Подобным же образом единица, расположенная в ярко освещенном поле поблизости от границы, будет иметь более высокую частоту разряда, чем другие одинаково освещенные единицы, расположенные также в ярком поле, но испытывающие более сильное торможение, так как их непосредственными соседями являются столь же ярко освещенные единицы. Таким образом, различия в активности элементов по обеим сторонам границы будут усилены и нарушение непрерывности освещения будет подчеркнуто в характере нервной реакции. Идеальной экспериментальной проверкой этих качественных предсказаний была бы одновременная регистрация разряда импульсов от большого числа рецепторных единиц во многих различных положениях, соответствующих фиксированному распределению освещенности на рецепторной мозаике Поскольку такая проверка неосуществима, измеряли разряд импульсов только от одной рецепторной единицы, расположенной поблизости от центра глаза, и в промежутках между измерениями изменяли освещение глаза таким образом, что данная рецепторная единица занимала последовательно ряд различных положений по отношению к участкам с разной освещенностью. Применяли две простые системы освещения: резкий скачок в интенсивности и простой градиент между двумя уровнями интенсивности. В каждом случае переходы в распределении освещенности подчеркиваются в соответствующем характере нервной реакции, а именно в частоте разряда рецептора появляются максимумы и минимумы на сторонах перехода освещенностей. Основное свойство большинства рецепторов заключается в том, что они не только сообщают сведения об устойчивом состоянии раздражителя, но также энергично реагируют на изменения интенсивности раздражителя во времени. Как для возбуждающего, так и для тормозного компонента активности сложного глаза Limulus характерны резкие кратковременные реакции на изменения раздражителя Любое изменение интенсивности освещенности вызывает значительное преходящее изменение частоты разряда, гораздо большее, чем соответствующее различие в частоте при разных стабильных состояниях. По- 15-1015 226 Глава IV Психофизиология зрительного восприятия 3 Сетчатка 227
Эти характерные преходящие режимы возбуждения оказывают сходное, но обратное влияние на частоту разрядов соседних элементов, возбуждение которых не изменялось. Следовательно, если сравнивать частоту разряда данного элемента с частотой разряда соседних элементов, преходящие частоты возбуждения будут казаться относительно небольшими, так как они вызывают обратный эффект в соседних элементах. Иногда, правда, после торможения наблюдается некоторое усиление реакции, несколько превышающее уровень незаторможенной реакции. Однако такое усиление всегда крайне мало. Преходящие процессы торможения протекают параллельно проходящим процессам возбуждения, но первые, вероятно, не полностью зависят от последних. Тормозное взаимодействие среди нервных элементов представляет собой один из главных нервных механизмов, общий для многих сенсорных модальностей и уровней интеграции. Основные функциональные свойства этого взаимодействия, по-видимому, одинаковы, где бы они не обнаруживались. Глаз Limulus особенно удобен для количественного анализа этих функциональных свойств: взаимодействие является чисто тормозным, группа взаимодействующих элементов относительно малочисленна и распределение возбуждения на рецепторной мозаике можно регулировать с большой точностью. Эксперименты с этим глазом моделируют относительно простой интегративный процесс, что может быть полезным для понимания более сложных интег-ративных процессов в других частях нервной системы, а также у животных других видов. В глазе Limulus четко выявляется влияние тормозного взаимодействия среди нервных элементов. Разряд импульсов в любом волокне зрительного нерва зависит не только от параметров раздражителя специфической рецепторной единицы, от которой отходит это волокно, но и от пространственного и временного распределения раздражения по всей популяции взаимодействующих элементов. Эти взаимодействия усиливают контраст при резких пространственных и временных градиентах и при нарушениях непрерывности изображения на сетчатке: края и контуры в их нервном представительстве становятся более четкими. Таким образом, возникающая активность зрительного нерва не является прямой копией распределения возбуждения на рецепторной мозаике. Определенная информация, особенно важная для организма, усиливается за счет менее важной информации. В сетчатке позвоночных взаимодействие протекает более сложно, что приводит к | большему разнообразию активности зрительного нерва. Одно влия-[ ние часто затемняет другое. Тем не менее, в зрительной системе позвоночных можно выделить отдельно участие возбуждающих и тормозных влияний на характер активности зрительного нерва. Например, Хартлайн обнаружил в глазе лягушки, что эффект выключения, вызванный в одиночном волокне посредством освещения одной группы рецепторов, можно затормозить, освещая другую группу рецепторов того же самого рецептивного поля. Установлено, что свет, совершенно не освещающий рецептивное поле данного волокна, но падающий поблизости от него, также оказывает тормозное влияние на реакции этого волокна. В глазе кошки определенные области в пределах одного рецепторного поля дают преимущественно или эффекты включения, или эффекты выключения. Когда в пределах одного рецептивного поля взаимодействуют две области с противоположными эффектами, реакции обеих, пройдя через их общий нейрон, изменяются. Дальнейшие изменения характера нервной активности происходят на высших интегративных уровнях зрительного пути. Например, тормозное взаимодействие нейронов обнаружено в зрительной коре кошки. Тормозное взаимодействие, несомненно, представляет собой основу ряда хорошо известных зрительных явлений, таких, как световой и цветовой контрасты. Действительно, ранние психофизические исследования этих явлений предопределили открытие тормозного взаимодействия физиологическими методами. В любом отдельном волокне зрительного нерва глаза Limulus, хирургически изолированном, можно возбудить активность только путем освещения той рецепторной единицы, от которой это волокно отходит (в глазе Limulus нет конвергенции возбуждения). Тем не менее на активность рецепторной единицы влияет освещение других, соседних, рецепторных единиц. Это влияние чисто тормозное. Активность каждого рецептора представляет собой результат действия на него светового раздражителя и торможения со стороны других рецепторов. При стабильном освещении торможение, оказываемое одной рецепторной единицей на другую, является линейной функцией частоты разряда первой единицы. Когда на данную рецепторную единицу действует несколько единиц, расположенных поблизости, их общее тормозное влияние количественно определяется суммой тормозных влияний каждой единицы. В результате реакции n-рецепторов, взаимодействующих друг с другом, можно описать посредством системы и-уравне-ний, линейных по частоте разрядов взаимодействующих единиц. Тормозное взаимодействие двух рецепторов глаза Limulus тем сильнее, чем ближе расположены они друг к другу. Этот фактор ока- 15* 228 Глава IV Психофизиология зрительно! о восприятия 4 Топографическое отображение 229
Кратковременные динамические фазы тормозного взаимодействия усиливают реакцию зрительного нерва на изменения интенсивности освещения. Действительно, при соответствующем столкновении возбуждающих влияний светового раздражения рецепторной единицы с тормозными влияниями от соседних рецепторов могут возникать кратковременные разряды импульсов, весьма напоминающие эффекты включения-выключения и эффекты выключения, столь характерные для различных волокон зрительного нерва сетчатки позвоночных. Эксперименты на глазе Limulus раскрывают один из механизмов, который, возможно, участвует в создании эффектов пространственного и временного контраста в более высокоорганизованных зрительных системах и, возможно, применим для расшифровки сложных интегра-тивных процессов в других частях нервной систем. 4. Топографическое отображение Постепенно были собраны сведения об участках мозга, связанных со зрительной функцией: достаточно хорошо прослежены начальные этапы передачи зрительной информации. Было установлено, что волокна зрительного нерва образуют синапсы с клетками наружного коленчатого тела (НКТ) и что аксоны клеток НКТ оканчиваются в первичной зрительной коре. Было также установлено, что эти связи — от сетчатки к НКТ и от НКТ к коре — имеют топографическую организацию. Говоря о топографическом отображении, мы имеем в виду, что предшествующая структура проецируется на последующую упорядоченным образом: если идти вдоль какой-либо линии на сетчатке, то проекции последовательных точек этой линии в НКТ и в коре также образуют одну непрерывную линию. Таким образом, волокна зрительного нерва, выходящие из небольшого участка сетчатки, все будут направляться к какому-то небольшому участку НКТ, а все волокна от небольшой зоны НКТ придут в определенную зону зрительной коры. Такая организация связей не покажется удивительной, если вспомнить упрощенную схему нервной системы, клетки группируются в структуру, напоминающую стопку пластин, причем каждая клетка любой пластины получает входы от некоторой компактной группы клеток предыдущей пластины. В сетчатке последовательные слои клеток расположены так, что нервные волокна могут кратчайшим путем проходить с одного уровня на следующий. Клетки наружных коленчатых тел удалены на некоторое расстояние от клеток сетчатки, точно так же как кора удалена от НКТ и находится в другой части мозга. Тем не менее характер соединения ее с клетками остается тем же — проекции отдельных зон на более высокие уровни организованы так, как если бы эти зоны точно накладывались одна на другую. На выходе из глазного яблока волокна зрительного нерва просто собираются в пучок. Дойдя до НКТ, они расходятся и образуют своими окончаниями упорядоченную проекцию. Удивительно то, что в зрительном нерве на пути от сетчатки к НКТ эти волокна почти полностью перепутываются, но на уровне НКТ снова находят свои места. Точно так же и волокна, которые выходят из НКТ, расходятся в виде широкой полосы, которая идет через весь мозг к затылку и оканчивается в первичной зрительной коре, где проекция снова оказывается упорядоченной. После того как эти пути, пройдя через первичную зрительную кору и образовав синапсы в различных ее слоях, выходят из этой области и достигают других корковых зон, они опять образуют топографически упорядоченную проекцию. Поскольку на каждом уровне есть конвергенция связей, рецептивные поля в целом постепенно становятся все ближе. Поэтому чем дальше от сетчатки, тем более размытым будет отображение зрительно воспринимаемой картины. Еще одним важным и давним свидетельством в пользу топографической организации зрительных путей служат клинические наблюдения. Если поврежден определенный участок первичной зрительной коры, то развивается «локальная слепота», как если бы была разрушена соответствующая область сетчатки. Есть определенная система в отображении зрительного мира в структурах наружных коленчатых тел и коры. Однако в 50-е гг. было неясно, что может означать такое отображение. В то время было еще слишком мало известно о закономерностях переработки информации на каждом из уровней ее анализа и интеграции, почти ничего не знали исследователи и о том, каков конечный результат переработки зрительных сигналов на каждом из уровней. Полагали, что картинка, созданная при помощи зрения, просто передается в мозг, а уж его задача — осмыслить ее. В дальнейшем стало понятно, что такая нейронная структура, как первичная зрительная кора, производит глубокое преобразование поступающей информации. Нам почти ничего не известно о том, что происходит на дальнейших этапах ее переработки. Но знание того, что определенная часть коры использует в своей работе
230 Глава IV Психофизиология зрительного восприятия 5. Зрительные области мозга 231
5. Зрительные области мозга Нервная система, ответственная за зрение, начинается с сетчатки. Сетчатка в сущности является вынесенным на периферию кусочком мозга, содержащим как типичные мозговые клетки, так и специализированные светочувствительные детекторы. Сетчатка делится по вертикали на две части; от наружных отделов сетчатки волокна идут к той же стороне затылочной области мозга, в то время как волокна от внутренней, назальной стороны сетчатки перекрещиваются сразу позади глаз — в хиазме (зрительный перекрест) и направляются к затылочной области противоположного полушария. Это зрительная область коры головного мозга. Мозг как целое делится на два полушария, каждое из которых представляет собой более или менее цельный мозг; оба полушария соединены массивным пучком волокон — corpus collossum (мозолистое тело). Волокна зрительного тракта от хиазмы идут в переключающие ядра каждого полушария, в область, называемую наружное коленчатое тело. У животных с фронтальным расположением глаз с темпоральной области часть волокон переходит на противоположную сторону. Этот процесс, по-видимому, проходит параллельно с развитием бинокулярного зрения. В этом и объяснение того, почему животные с фронтальным расположением глаз могут видеть височной частью сетчатки то же, что видит другой глаз назальной сетчаткой. Наружное коленчатое тело Волокна, идущие в мозг от каждого глаза, проходят через зрительную хиазму. В хиазме примерно половина волокон каждого зрительного нерва переходит на противоположную сторону мозга по отношению к данному глазу, а другая половина остается на той же стороне мозга. Пройдя хиазму, волокна направляются в несколько разных пунктов. Некоторые из них идут в нейронные структуры, имеющие отношение к таким специфическим реакциям, как движения глаз и зрачковый рефлекс. Однако большая часть волокон оканчивается в двух наружных коленчатых телах (НКТ) (рис. 14). По сравнению с корой и множеством других отделов мозга эти тела устроены сравнительно просто — все или почти все из примерно полутора миллионов клеток в каждом НКТ имеют непосредственные входы от волокон зрительного нерва, и большинство клеток (но не все) посылают свои
|
||||||||||
|
|
Последнее изменение этой страницы: 2021-01-09; просмотров: 95; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.58.143 (0.087 с.) |




 нальной передачей. Ясно, что рецепторы доставляют информацию горизонтальным клеткам через возбуждающие синапсы, поскольку в большинстве случаев горизонтальные клетки, подобно рецепторам, гиперполяризуются (выключаются) светом. Не совсем понятно, куда эти клетки посылают свои выходные сигналы. Известно, что у некоторых животных, например, у черепах, они передают информацию обратно рецепторам; у других видов они образуют синапсы с дендрита-ми биполярных клеток и, несомненно, передают им информацию; о приматах нет каких-либо конкретных сведений. Горизонтальные клетки получают входные сигналы от рецепторов; их выход пока точно неизвестен, но он направлен или к рецепторам, или к биполярным клеткам, или к тем и другим.
нальной передачей. Ясно, что рецепторы доставляют информацию горизонтальным клеткам через возбуждающие синапсы, поскольку в большинстве случаев горизонтальные клетки, подобно рецепторам, гиперполяризуются (выключаются) светом. Не совсем понятно, куда эти клетки посылают свои выходные сигналы. Известно, что у некоторых животных, например, у черепах, они передают информацию обратно рецепторам; у других видов они образуют синапсы с дендрита-ми биполярных клеток и, несомненно, передают им информацию; о приматах нет каких-либо конкретных сведений. Горизонтальные клетки получают входные сигналы от рецепторов; их выход пока точно неизвестен, но он направлен или к рецепторам, или к биполярным клеткам, или к тем и другим.





 представить себе нервы с частотой разрядов, поддерживаемой на достаточно высоком уровне, скажем, 100 в с, и потому способных в ответ на оппонентные стимулы либо снижать, либо повышать частоту импульсов — вниз до нуля, а вверх, например, до 500 в с. Но поскольку на импульсы расходуется метаболическая энергия (весь входящий в нерв натрий должен откачиваться наружу), то для нервных клеток в состоянии покоя более экономным, вероятно, будет молчание импуль-сации с низкой частотой, а организму выгодно иметь для каждой данной модальности две отдельные группы клеток, одна из которых разряжается при уменьшении, а другая при увеличении силы стимула.
представить себе нервы с частотой разрядов, поддерживаемой на достаточно высоком уровне, скажем, 100 в с, и потому способных в ответ на оппонентные стимулы либо снижать, либо повышать частоту импульсов — вниз до нуля, а вверх, например, до 500 в с. Но поскольку на импульсы расходуется метаболическая энергия (весь входящий в нерв натрий должен откачиваться наружу), то для нервных клеток в состоянии покоя более экономным, вероятно, будет молчание импуль-сации с низкой частотой, а организму выгодно иметь для каждой данной модальности две отдельные группы клеток, одна из которых разряжается при уменьшении, а другая при увеличении силы стимула. От греч omma — глаз — элемент сложного глаза насекомого и членистоногих
От греч omma — глаз — элемент сложного глаза насекомого и членистоногих



 освещении соседних омматидиев, а именно: увеличивается порог светового раздражителя, уменьшается число импульсов, возникающих в ответ на надпороговую вспышку света, и понижается частота разряда импульсов при стабильном освещении. Показано, что степень торможения, о которой судят по понижению частоты разряда, зависит от интенсивности освещения, от площади и конфигурации изображения на сетчатке: 1) чем больше интенсивность освещения соседних рецепторов, тем сильнее тормозное действие, оказываемое ими на исследуемый рецептор; 2) чем больше число освещенных соседних рецепторов, т.е. чем больше площадь освещения, тем сильнее торможение исследуемого рецептора; 3) освещение рецепторов, расположенных в непосредственной близости к исследуемому рецептору, приводит к более сильному торможению, чем освещение удаленных рецепторов.
освещении соседних омматидиев, а именно: увеличивается порог светового раздражителя, уменьшается число импульсов, возникающих в ответ на надпороговую вспышку света, и понижается частота разряда импульсов при стабильном освещении. Показано, что степень торможения, о которой судят по понижению частоты разряда, зависит от интенсивности освещения, от площади и конфигурации изображения на сетчатке: 1) чем больше интенсивность освещения соседних рецепторов, тем сильнее тормозное действие, оказываемое ими на исследуемый рецептор; 2) чем больше число освещенных соседних рецепторов, т.е. чем больше площадь освещения, тем сильнее торможение исследуемого рецептора; 3) освещение рецепторов, расположенных в непосредственной близости к исследуемому рецептору, приводит к более сильному торможению, чем освещение удаленных рецепторов.



 добные преходящие изменения, по-видимому, зависят как от скорости изменения освещенности, так и от абсолютной величины медленных «генераторных» потенциалов в омматидии.
добные преходящие изменения, по-видимому, зависят как от скорости изменения освещенности, так и от абсолютной величины медленных «генераторных» потенциалов в омматидии. зывает важное влияние на активность зрительного нерва, возбуждаемую различиями в распределении освещенности на рецепторной мозаике: происходит усиление контраста в области резких градиентов изображения на сетчатке. Эти эффекты весьма напоминают аналогичные эффекты в зрении человека (полосы Маха и пограничный контраст).
зывает важное влияние на активность зрительного нерва, возбуждаемую различиями в распределении освещенности на рецепторной мозаике: происходит усиление контраста в области резких градиентов изображения на сетчатке. Эти эффекты весьма напоминают аналогичные эффекты в зрении человека (полосы Маха и пограничный контраст).




 вполне понятные принципы, дает основание для предположении о том, что и остальные зоны коры работают так же.
вполне понятные принципы, дает основание для предположении о том, что и остальные зоны коры работают так же.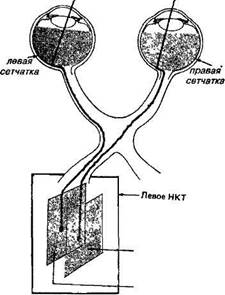 аксоны в кору мозга. Отсюда следует, что пути, проходящие через НКТ в кору, имеют только одно синаптическое переключение; Однако было бы ошибкой считать НКТ просто передаточной станцией. Сюда входят не только волокна из зрительного нерва, но и волокна, приходящие обратно из тех участков коры, на которые проецируются НКТ, а также из ретикулярной формации ствола мозга, имеющей отношение к процессам внимания и общей активации. Некоторые клетки НКТ имеют короткие аксоны (меньше мм длиной), они не выходят за пределы НКТ, а образуют синаптические контакты с другими нейронами НКТ. Несмотря на эти усложнения, одиночные клетки НКТ отвечают на световые стимулы примерно так же, как и ганглиозные клетки сетчатки, и у них сходная структура рецептивных полей с on - и о//-центрами и сходные ответы на цветовые стимулы. Таким образом, если говорить о переработке зрительной информации, НКТ, по-видимому, не производит никаких значительных преобразований сигналов. Что же касается незрительных входов НКТ и локальных синап-тических связей, то пока мы просто ничего не знаем об их роли.
аксоны в кору мозга. Отсюда следует, что пути, проходящие через НКТ в кору, имеют только одно синаптическое переключение; Однако было бы ошибкой считать НКТ просто передаточной станцией. Сюда входят не только волокна из зрительного нерва, но и волокна, приходящие обратно из тех участков коры, на которые проецируются НКТ, а также из ретикулярной формации ствола мозга, имеющей отношение к процессам внимания и общей активации. Некоторые клетки НКТ имеют короткие аксоны (меньше мм длиной), они не выходят за пределы НКТ, а образуют синаптические контакты с другими нейронами НКТ. Несмотря на эти усложнения, одиночные клетки НКТ отвечают на световые стимулы примерно так же, как и ганглиозные клетки сетчатки, и у них сходная структура рецептивных полей с on - и о//-центрами и сходные ответы на цветовые стимулы. Таким образом, если говорить о переработке зрительной информации, НКТ, по-видимому, не производит никаких значительных преобразований сигналов. Что же касается незрительных входов НКТ и локальных синап-тических связей, то пока мы просто ничего не знаем об их роли.


