Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Опыты над гибридами других видов растений
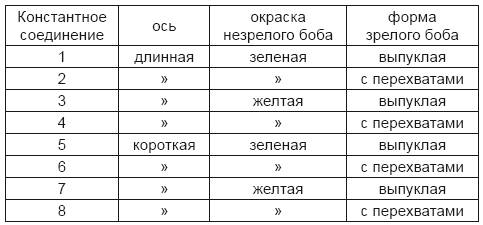
Задачей дальнейших опытов было определить, приложим ли и к гибридам других растений закон развития, найденный для гороха. С этой целью в последнее время были поставлены многочисленные опыты. Закончены два небольших опыта с видами Phaseolus, которые и могут быть упомянуты здесь. Опыт с Phaseolus vulgaris и Phaseolus nanus дал вполне согласный результат. Ph. nanus имеет наряду с карликовыми осями зеленые просто выпуклые бобы; Ph. vulgaris, наоборот, – оси длиною 10–12 и желтые бобы с перехватами ко времени созревания. Числовые отношения, в которых появлялись различные формы в отдельных поколениях, были такими же, как и у Pisurn. Так же и развитие константных соединений следовало закону простого комбинирования признаков, точно так, как это происходит у Pisurn. Было получено:
Зеленая окраска боба, выпуклая форма боба и длинная ось были, как и у Pisurn, доминирующими признаками. Другой опыт с двумя очень различающимися видами Phaseolus имел только частичный успех. Семенным растением служил Phaseolus nanus, вполне константный вид с белыми цветами в коротких кистях и маленькими белыми семенами в прямых, выпуклых и гладких бобах; пыльцевым растением был Ph. multiflorus с высоко вьющимся стеблем, пурпурно-красными цветами в очень длинных кистях, с опушенными серповидно искривленными бобами и большими семенами с черными пятнами на персиковом кроваво-красном фоне. Гибрид очень походил на пыльцевое растение, только цветы были менее интенсивно окрашены. Плодовитость их была очень ограниченна: от 17 растений, которые дали вместе многие сотни цветов, было собрано в общем только 49 семян. Они были средней величины и имели рисунок, одинаковый с Ph. multiflorus; основная окраска также не отличалась ничем существенным. В следующем году от них было получено 44 растения, из которых только 31 достигло цветения. Признаки от Ph. nanus, которые в гибриде были целиком патентны, снова обнаружились в различных комбинациях; отношение их к доминирующим вследствие незначительного числа опытных растений было колеблющимся; для некоторых признаков, как для оси и для формы боба, отношение было, как у Pisurn, почти точно 1:3. Как ни мал успех этого опыта для установления числовых отношений, в которых появляются различные формы, но, с другой стороны, этот опыт представляет собою случай замечательного изменения окраски цветов и семян у гибридов. У Pisurn, как известно, признаки окраски цветов и семян появляются неизменными в первом и последующих поколениях и потомки гибридов несут исключительна тот или другой из двух исходных признаков. Иначе обстоит дело в настоящем опыте. Белая окраска цветов и семян Ph. nanus действительно появилась уже в первом поколении, но лишь на одном плодовитом экземпляре, у остальных же 30 растений окраска цветов представляла различные переходы от пурпурно-красного до бледно-фиолетового. Окраска семенной кожуры была не менее разнообразна, чем у цветов. Ни одно растение нельзя было считать вполне плодовитым; некоторые растения совсем не завязывают плодов, у других плоды развиваются только из последних цветов и не достигают созревания; только с 15 растений были собраны хорошо развившиеся семена. Наибольшую склонность к бесплодию обнаруживали формы с преобладающими красными цветами, из них 16 растений дали только 4 зрелых семени. У трех из них был одинаковый с Ph. multiflorus рисунок семян, однако с более или менее бледным основным фоном, четвертое растение дало только одно семя с однообразно коричневой окраской, формы с преобладающей лиловой окраской цветов имели темно-коричневые, черно-коричневые и совсем черные семена.
Опыт был проведен еще через два поколения при таких же неблагоприятных обстоятельствах, так как даже потомки плодовитых растений были частью менее плодовиты, частью совсем бесплодны. Других окрасок семян и цветов, кроме вышеописанных, больше не появлялось. Формы, которые в первом поколении получали один или несколько рецессивных признаков, оставались по отношению к этим признакам все без исключения константными. Некоторые из тех растений, которые имели лиловые цветы и коричневые или черные семена, в следующих поколениях тоже не изменили больше окраски цветов и семян, однако большинство из них дало наряду с совершенно одинаковыми потомками такие, которые получили белые цветы и так же окрашенную кожуру семян. Растения с красными цветами оставались столь мало плодовитыми, что об их дальнейшем развитии нельзя сказать ничего определенного.
Несмотря на многочисленные помехи, с которыми приходилось бороться при наблюдении, все же из этого опыта ясно, что развитие гибридов по отношению к тем признакам, которые касаются формы растения, следует тем же законам, как и у Pisurn. Относительно признаков окраски затруднительно обнаружить достаточную согласованность. Кроме того, что из соединения белой и пурпурно-красной окраски получается целый ряд оттенков, от пурпурного до бледно-лилового и белого, бросается в глаза еще то обстоятельство, что между 31 цветущими растениями только одно получило рецессивный признак белой окраски, тогда как у Pisurn он появляется в среднем у каждого четвертого растения. Но также и эти загадочные явления можно было бы правдоподобно объяснить законами, установленными на Pisurn, если предположить, что окраска цветов и семян Ph. multiflorus состоит из двух или нескольких совершенно самостоятельных окрасок, которые отдельно ведут себя совершенно так же, как любой другой константный признак у растения. Если окраска цветов А слагается из самостоятельных признаков А1 + А2 +…, которые вызывают общее впечатление пурпурно-красной окраски, то при оплодотворении формы с различающимся признаком белой окраски а должны образовываться гибридные соединения А1а + А2а +…; то же самое происходит с соответствующей окраской семенной кожуры. При вышеупомянутом предположении каждое из этих гибридных соединений окраски самостоятельно и развивается поэтому совершенно независимо от остальных. Отсюда легко видеть, что из комбинации отдельных рядов развития может получиться полный ряд окрасок. Если, например, А = А1 + А2, то гибриды А1а и А2а соответствуют рядам А1 + 2A1a + a А2 + 2А2a + a. Члены этих рядов могут войти в 9 различных соединений, и каждое из них представляет обозначение для особой окраски:
Числа отдельных соединений показывают, сколько растений с соответственной окраской принадлежит данному ряду. Так как сумма этого последнего равняется 16, то все окраски в среднем распределяются между каждыми 16 растениями, однако, как показывает сам ряд, в неодинаковых отношениях. Если развитие окраски происходит действительно таким образом, то может найти себе объяснение и тот вышеприведенный случай, когда белая окраска цветов и бобов встретилась среди 31 растения первого поколения только один раз. Эта окраска содержится в ряду только однажды, и поэтому может развиться в среднем по одному разу на каждые 16, а при трех признаках окраски даже на каждые 64 растения. Однако не следует забывать, что приведенное здесь объяснение основывается на одном лишь предположении, которое ничего больше, кроме очень неполного результата вышеизложенного опыта, не имеет за собой. Но если бы проследить развитие окраски гибридов в таких опытах дальше, то это была бы многообещающая работа, так как, вероятно, мы могли бы научиться понимать таким путем необычайное разнообразие окраски наших декоративных цветов. До сих пор едва ли достоверно известно больше того, что окраска цветов у большинства декоративных растений является крайне изменчивым признаком. Часто высказывалось мнение, что устойчивость видов оказывается сильно поколебленной или совсем разрушается культивированием, и существует сильная склонность представлять развитие культурных форм неправильным и случайным; при этом обычно указывается на окраску декоративных растений как на пример непостоянства. Однако нельзя постигнуть, почему одно перемещение на садовую почву должно производить столь глубокую и продолжительную революцию в растительном организме. Никто не станет серьезно утверждать, что развитие на воле управляется другими законами, чем на садовой грядке. Как здесь, так и там должны появиться характерные изменения, если для данного вида изменяются жизненные условия и этот вид обладает способностью приспособляться к новым обстоятельствам. Вполне допустимо, что культура благоприятствует возникновению новых разновидностей, и человек сохраняет многие изменения, которые не удержались бы на свободе; все же это не дает нам основания предполагать, что склонность к образованию разновидностей столь необычайно повышается, что виды теряют всякую самостоятельность, а их потомки расходятся в бесконечные ряды высокоизменчивых форм. Если бы изменение в условиях произрастания было единственной причиной изменчивости, то следовало бы ожидать, что те культурные растения, которые в течение столетий возделываются при почти одинаковых условиях, должны были вновь приобрести известную устойчивость. Как известно, этого не наблюдается, так как именно между ними мы находим не только самые разнообразные, но также и самые изменчивые формы. Только бобовые, как Pisurn, Phaseolus, Lens, половые органы которых закрыты лодочкой, представляют замечательное исключение. Хотя и у них в течение больше чем 1000-летней культуры при разнообразных условиях возникли многочисленные разновидности, но в одинаковых условиях существования они удерживают такую же самостоятельность, какая свойственна дикорастущим видам.
Более чем вероятно, что в изменчивости культурных растений действует фактор, которому до сих пор уделяли мало внимания. Различные наблюдения и опыты (Erfahrungen) заставляют полагать, что наши культурные растения за немногими исключениями являются членами различных гибридных рядов, дальнейшее закономерное развитие которых нарушается и задерживается частыми взаимными скрещиваниями. Не следует упускать из виду, что культурные растения большей частью выращиваются в больших количествах совместно, и этим предоставляется благоприятная возможность для взаимного оплодотворения между разновидностями и даже видами. Вероятность этого взгляда поддерживается тем фактом, что среди множества изменчивых форм всегда находятся некоторые остающиеся в том или ином признаке константными, если только тщательно охранять их от всякого чужого влияния. Эти формы размножаются точно так же, как определенные члены сложных гибридных рядов. Даже по отношению к самому чувствительному из всех признаков, к окраске, при внимательном наблюдении не ускользает, что у отдельных форм наклонность к изменчивости проявляется в очень различной степени. Между растениями, которые происходят в результате одного и того же произвольного оплодотворения, часто встречаются такие, потомки которых далеко отстоят друг от друга в особенностях и распределении окраски, тогда как другие дают мало уклоняющиеся формы, и среди множества растений попадаются такие, которые передают потомкам свою окраску цветов неизменной. Культивируемые виды Dianthus дают тому поучительное доказательство. Цветущий белыми цветами экземпляр Dianthus caryophyllus, который сам произошел от разновидности с белыми цветами, был перенесен во время цветения в теплицу; полученные от него многочисленные семена дали растения с совершенно одинаковой белой окраской цветов. Сходный результат был получен от красного, немного отливающего лиловым и от белого с красными полосами подвида. Наоборот, многие другие подвиды, таким же образом защищенные, дали более или менее различно окрашенных и разрисованных потомков.
Кто обозревает окраски, которые появляются у декоративных растений из одного и того же оплодотворения, с трудом может отказаться от убеждения, что и здесь развитие подчиняется также определенному закону, выражающемуся, по-видимому, в комбинировании многих самостоятельных признаков окраски.
Заключительные замечания
Небезынтересно сравнить наблюдения, сделанные у Pisurn, с результатами, к которым пришли в своих исследованиях два авторитета в этой области – Кельрейтер и Гэртнер. По их согласному представлению, гибриды по своим внешним проявлениям или держатся средней формы между обоими исходными видами, или же они подходят ближе к типу того или другого из них, иногда едва отличаясь от него. Из этих семян, если происходит опыление собственной пыльцой, обыкновенно выходят различные формы, отклоняющиеся от нормального типа. Как правило, большинство индивидов из такого оплодотворения удерживает форму гибрида, тогда как немногие другие растения больше похожи на семенное растение и один-два индивида близко подходят к пыльцевому растению. Однако это не относится ко всем гибридам без исключения. В некоторых случаях потомки стоят ближе то к одному, то к другому исходному растению или все они склоняются больше в одну какую-нибудь сторону, но у некоторых [растений] они остаются совершенно подобными, гибридами размножаются дальше, не изменяясь. Гибриды разновидностей ведут себя, как гибриды видов, только обладают еще большей изменчивостью формы и еще большей склонностью возвращаться к исходным формам.
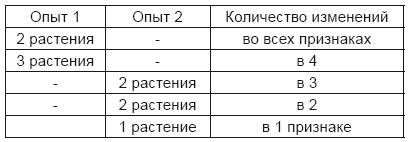
В отношении формы гибридов и их обычного развития нельзя не признать известного сходства с наблюдениями, сделанными над Pisurn. Иначе обстоит дело с упомянутыми исключительными случаями. Гэртнер признается сам, что точное определение, на какой из двух исходных видов больше похожа данная форма, встречает большие затруднения, так как здесь надо отнести многое на счет субъективного впечатления наблюдателя. Затем здесь следует принять во внимание еще одно обстоятельство, а именно, что, несмотря на тщательное наблюдение и различение, результаты были неустойчивые и неопределенные. Для опытов служили большей частью растения, которые считались хорошими видами и отличались во многих признаках. Наряду с резко выступающими признаками, там, где приходилось считаться с большим или меньшим общим сходством, нужно было обращать внимание и на такие признаки, которые трудно определить словами, но которых, тем не менее, достаточно, чтобы придать формам другой вид, как это известно каждому знатоку растений. Если признать, что развитие гибридов следует закону, установленному для Pisurn, то ряд в каждом отдельном опыте должен охватить очень много форм, так как число членов, как известно, увеличивается в третьей степени по отношению к числу различающихся признаков. При относительно незначительном числе опытных растений результат должен быть только приблизительно правильным и в отдельных случаях может значительно отклоняться. Если, например, два исходных вида различаются в семи признаках и из семян их гибридов выращено от 100 до 200 растений для установления степени родства потомства, то нам ясно, как шатко будет суждение на основании такого материала, так как для семи различающихся признаков ряд в 16 384 индивида содержит 2187 различных форм. В зависимости от того, какая форма попадет случайно в руки наблюдателя в большем количестве, может быть придано большее значение то одному, то другому сродству. Далее, если среди различающихся признаков встречаются доминирующие, которые переходят к гибриду совершенно или почти совершенно неизмененными, то среди членов ряда будет выступать на первый план тот из обоих исходных видов, который обладает большим числом доминирующих признаков. В опыте с Pisurn, приведенном раньше для трояко различающихся признаков, доминирующие признаки принадлежали все семенному растению, и хотя по своему внутреннему составу члены ряда склонялись равномерно к обоим исходным растениям, все же тип семенного растения получил в этом опыте такой значительный перевес, что между каждыми 64 растениями первого поколения 54 были совершенно одинаковы с ним или отличались от него только одним признаком. Отсюда видно, как рискованно при некоторых обстоятельствах по внешнему сходству гибридов делать заключения об их внутреннем сродстве. Гэртнер упоминает, что в тех случаях, когда развитие идет закономерно, среди потомков гибридов получаются не сами исходные виды, но только единичные особи, близко стоящие к ним. При очень обширном ряде развития ничего другого нельзя было и ожидать. Например, для семи различающихся признаков среди 16 000 потомков гибридов обе исходные формы встречаются только по одному разу. Поэтому получить их уже при небольшом числе опытных растений мало возможно, однако с некоторой вероятностью можно рассчитывать на появление таких отдельных форм, которые стоят в ряду близко к исходным видам. Существенное отличие мы встречаем у тех гибридов, которые остаются константными в потомстве и размножаются как чистые виды. По Гэртнеру, сюда принадлежат превосходно плодовитые гибриды: Aquilegia atropurpurea-canadensis, Lavatera pseudolbia-thuringiaca, Geum urbanorivale и некоторые гибриды Dianthus; по Вихура – гибриды видов ив. Для истории развития растений это обстоятельство имеет особенное значение, так как константные гибриды получают значение новых видов. За правильность этого обстоятельства ручается имя превосходного наблюдателя, и оно не может подвергаться сомнению. Гэртнер имел возможность проследить Dianthus Armeria-deltoidcs до 10 поколения, так как это растение нормально размножалось в саду само по себе в одну единую клетку, которая развивается в самостоятельный организм путем восприятия веществ и образования новых клеток. Это развитие следует постоянному закону, который основывается на материальном составе и расположении элементов, достигших в клетке жизнеспособного соединения. Если воспроизводительные клетки однородны и подобны основной клетке материнского растения, тогда развитие нового индивида управляется тем явнее законом, которому подчиняется материнское растение. Если же удается соединить зачатковую клетку с неоднородной пыльцевой клеткой, то мы должны признать, что между теми элементами обеих клеток, которые обусловливают взаимные различия, осуществляется некое уравновешивание (Ausgleichung). Образующаяся из них некоторая согласованная клетка (Vermittlungszellе) является основой для гибридного организма, развитие которого обязательно следует другому закону, чем у каждого из обоих исходных видов. Если признать уравновешивание полным в том смысле, что гибридный зародыш образуется из одинаковых клеток, в которых различия совершенно и прочно согласованы, то надо признать дальнейшее следствие, что гибрид должен оставаться в своих потомках константным, как всякий другой самостоятельный вид растения. Воспроизводительные клетки, которые образуются в завязях и пыльниках такового, однородны и вполне сходны с основной согласованной клеткой. Относительно тех гибридов, потомки которых изменчивы, можно, по-видимому, признать, что между различающимися элементами зачатковой и пыльцевой клетки происходит согласование лишь в той мере, в какой это необходимо для образования клетки как основания для гибрида, но уравновешивание противоборствующих (widerstrebenden) элементов здесь только преходящее и не распространяется за пределы жизни гибридного растения. Так как во время всего периода вегетации во внешнем виде его нет никаких видимых изменений, то мы должны заключить, что различающимся между собою элементам удается выйти из вынужденного соединения только при развитии половых клеток. При образовании этих клеток все наличные элементы распределяются в совершенно свободных и равномерных группировках, и лишь различающиеся элементы при этом взаимно исключают друг друга. Таким путем возможно возникновение стольких зачатковых и пыльцевых клеток, сколько различных комбинаций допускают способные образоваться элементы. Сделанная здесь попытка свести существенное различие в развитии гибридов на постоянное или преходящее соединение различающихся клеточных элементов имеет, само собой разумеется, значение только гипотезы, в которой еще многое остается открытым из-за недостатка достоверных данных. Некоторое оправдание высказанному предположению заключается в доказательствах, приведенных для Pisurn, что поведение каждых двух различающихся признаков в гибридном соединении независимо от остальных различий между двумя исходными растениями, и затем, что гибриды производят столько зачатковых и пыльцевых клеток, сколько возможно константных комбинационных форм. Отличающиеся признаки двух растений могут зависеть в конце концов лишь от различий в свойствах и группировке элементов, которые находятся в жизненном взаимодействии в основных клетках этих растений. Правильность положений, установленных для Pisurn, сама еще нуждается, однако, в подтверждении, и поэтому желательно повторение по крайней мере наиболее важных опытов, например, относительно свойства гибридных половых клеток. От одного наблюдателя легко может ускользнуть какое-нибудь расхождение, которое покажется сначала незначительным, а потом может так возрасти, что им нельзя будет пренебрегать при установлении общего результата. Точно так же только опыт может решить, вполне ли сходно ведут себя изменчивые гибриды других видов растений; однако следует предполагать, что в основных моментах не может быть принципиального различия, так как единство плана развития органической жизни стоит вне сомнения. В заключение следует снова напомнить опыты, произведенные Кёльрейтером, Гэртнером и др. над превращением одного вида в другой путем искусственного оплодотворения. Этим опытам приписывалось особенное значение, и Гэртнер относит их к «труднейшим при получении бастардов». Если вид А должен быть превращен в другой вид В, то оба соединяются путем оплодотворения, и полученные гибриды еще раз оплодотворяются пыльцой В; затем из разнообразных потомков выбирается та форма, которая стоит ближе, и вновь оплодотворяется этой последней и так далее, пока не получится форма, одинаковая с В, которая и остается в своем потомстве константной. Таким образом, вид А превращается в другой вид В. Гэртнер один произвел 30 таких опытов с растениями из родов: Aquilegia, Dianthus, Geurn, Lavatera, Lychnis, Malvat Nicotiana и Onothera. Продолжительность превращения неодинакова для разных видов: тогда как для некоторых достаточно трехкратного оплодотворения, у других оно должно быть повторено от 5 до 6 раз, но и у одних и тех же видов наблюдались колебания в различных опытах. Гэртнер приписывает это различие тому обстоятельству, что «типовая сила, с которой определенный вид воспроизведения влияет на изменение и превращение материнского типа, крайне различна у различных растений, и следовательно, должны быть очень различны как периоды, в которые один вид превращается в другой, так и число поколений, нужных для этого, и превращение у одних видов происходит через многие, у других через небольшое число поколений». Дальше тот же наблюдатель замечает, «что и у продуктов превращения все зависит от того, какой тип и какой индивидуум будет выбран для дальнейшего превращения». Если позволено предполагать, что в подобных опытах развитие форм происходит таким же образом, как у Pisurn, тогда для всего процесса превращения можно найти довольно простое объяснение. Гибрид образует столько зачатковых клеток, сколько константных комбинаций допускают соединяющиеся в нем признаки, и одна из них всегда тождественна с оплодотворяющими пыльцевыми клетками. Поэтому во всех таких опытах всегда возможно, что уже из второго оплодотворения получается константная форма, которая подобна пыльцевому растению. Но будет ли получена в действительности эта последняя, зависит в каждом отдельном случае от числа опытных растений, а также от числа различающихся признаков, которые соединяются путем оплодотворения. Предположим, например, что растения, предназначенные для опыта, были различны в трех признаках и вид АВС должен превратиться в другой abc повторным оплодотворением пыльцой этого последнего типа. Гибрид, полученный из первого оплодотворения, образует 8 различных видов зачатковых клеток, а именно: АВС, АВс, АbС, аВС, Aba, аВс, abC, abc. Они соединяются во втором опытном году еще раз с пыльцевыми клетками abc, и получается ряд: АаВbСс + АаВbс + AabCc + аВbСс + Aabc + аВbс + аbСс + abc. Так как форма аbс в 8-членном ряду встречается один раз, то мало вероятно, что она могла бы отсутствовать даже в том случае, если растения выращивались в незначительном числе; превращение заканчивалось бы при этом уже после двукратного оплодотворения. Если бы превращение случайно не получалось, то надо было бы повторить скрещивание с одним из родственных соединений Ааbс, аВbс, аbСс. Ясно, что такой опыт должен длиться тем дольше, чем меньше число опытных растений и чем больше число различающихся признаков в обоих исходных видах; таким образом, у этих видов легко может произойти запаздывание на одно и даже два поколения, как это наблюдал Гэртнер. Превращение далеко отстоящих видов может закончиться все же лишь на 5-м или 6-м году опытов, так как число различных зачатковых клеток, которые образуются у гибрида, увеличивается с каждым лишним отличительным признаком пропорционально второй степени. Гэртнер путем повторных опытов нашел, что продолжительность взаимного превращения для некоторых видов различна, так что часто вид А превращается в другой В на одно поколение раньше, чем вид В в вид А. Отсюда он выводит заключение, что воззрение Кёльрейтера, по которому «оба естества у гибридов удерживают полнейшее равновесие», не совсем основательно. Однако возможно, что Кёльрейтер не заслуживает этого упрека; что скорее Гэртнер проглядел один важный момент, на который он сам обращает внимание в другом месте, а именно, что «это зависит от того, какой индивидуум был выбран для дальнейшего превращения». Опыты, которые были поставлены в этом направлении с двумя видами Pisurn, доказывают, что при выборе индивидов, более пригодных для целей дальнейшего оплодотворения, может быть большое различие от того, какой именно из двух видов превращается в другой. Оба опытных растения отличались пятью признаками, причем вид А имел все доминирующие, вид В – все рецессивные признаки. Для взаимного превращения А оплодотворялось пыльцой В и, обратно, В – пыльцой А, затем то же повторялось для обоих гибридов в следующем году. При первом опыте в третьем году имелось налицо 87 растений, в 32 возможных формах для выбора индивидов к дальнейшему оплодотворению; при втором опыте было получено 73 растения, которые по своему внешнему виду были вполне сходны с пыльцевым растением, однако их внутренние свойства были так же разнообразны, как формы в первом опыте. Рассчитанный выбор мог быть сделан только в первом опыте, во втором же те или иные растения могли быть выделены чисто случайно. Из последних только часть цветов была оплодотворена пыльцой А, другие предоставлены самооплодотворению. Из каждых 5 растений, выделенных в обоих опытах для оплодотворения, оказались сходными с пыльцевым растением, как показал сбор ближайшего года:
Для первого опыта, таким образом, превращение было закончено, для второго, который дальше не продолжался, необходимо было бы сделать вторичное оплодотворение. Если и не часто бывает такой случай, когда все доминирующие признаки принадлежат исключительно тому или другому основному растению, то все-таки всегда остается различие в том, которое из двух растений имеет их большее число. Если большинство доминирующих признаков принадлежит пыльцевому растению, тогда выбор форм для дальнейшего оплодотворения менее надежен, чем в обратном случае; а это ведет к замедлению во времени превращения, так как опыт можно будет считать законченным только тогда, когда получится форма, не только похожая на пыльцевое растение, но также остающаяся константной в потомстве, как и это последнее. Успех опытов с превращениями заставил Гэртнера высказаться против мнения тех естествоиспытателей, которые оспаривают постоянство видов растений и признают постоянное новообразование растительных видов. Он видит в окончательном превращении одного вида в другой недвусмысленное доказательство, что виду положены твердые границы, через которые он не может перейти. Если за этим взглядом нельзя признать безусловного значения, то, с другой стороны, в опытах, поставленных Гэртнером, находится достойное внимания подтверждение высказанного выше предположения об изменяемости культурных растений. Среди опытных видов были культурные растения, как Aquilegia atropurpurea и canadensis, Dianthus Caryophyllus, chinensis и japonicus, Nicotiana rnstica и paniculata, и они также не потеряли ничего в своей самостоятельности после 4– и 5-кратного гибридного оплодотворения.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 118; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.131.168 (0.025 с.) |