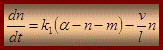Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Молекулярні механізми м”язового скорочення
В основі м'язового скорочення лежить взаємодія між товстою та тонкою протофібрилами, яка регулюється іонами Са2+. Запропонована стерична модель регуляції скорочення: Іони Са2+ зв'язуються з ТН-Са2+, викликаючи зміни в структурі білків тонкої нитки протофібрили через зміни ТМ і актин до АТФ-азного центру міозину. Якщо іони Са2+ в клітині містяться у концентрації 10-8 М, то ТМ знаходиться під кутом 50о до осі протофібрили, і субодиниця ТН-І не дозволяє взаємодіяти актину з міозином. При виникненні збудження ТН-Са2+ зв'язує відповідні іони, ТМ сповзає в борозну під кутом 70о, і ТН-І звільняє центр зв'язування на актин. МІОЗИН ТА АТФ-азна РЕАКЦІЯ Для виконання механічної роботи м'яз використовує енергію АТФ. Міозинову АТФазу відкрили Енгельгардт й Любимова (1939), кінетика цього ферменту не описується рівнянням Міхаеліса-Ментен. Цей фермент має певні особливості – здатні утворювати інтермедіати (проміжні конформації М* і М** багаті енергією). При взаємодії А-М з АТФ утворюється комплекс АМ**ФАДФ, якщо А заблокований, то М*+АТФ↔М**ФАДФ; Стадія АМ**ФАДФ↔АМ*ФАДФ значно швидша ніж ТЕОРІЯ М”ЯЗОВОГО СКОРОЧЕННЯ Першою була кінетична теорія Хакслі (модель взаємного ковзання двох типів протофібрил, 1954) – товсті та тонкі протофібрили ковзають одна відносно одної, не міняючи довжини, сила яку розвиває саркомер пропорційна кількості мостиків в зоні перекриття фібрил. Модель Дещеревського (1968) – на товстих протофібрилах є мостики, які замикаються на тонких протофібрилах. Існує три стани мостика: вільний; замкнутий, що тягне з силою f і замкнутий, що гальмує, з силою f. Виходячи з циклічної роботи мостика Дещеревський приходить до рівняння Хілла. ТЕОРІЯ М”ЯЗОВОГО СКОРОЧЕННЯ
де α – загальна кількість мостиків; n – тягнучі мостики; m – гальмуючі мостики; k1 – швидкість переходу з вільного стану до замкнуто тягучих; k2 - швидкість переходу з замкнуто тягучого стану до замкнуто гальмівного; l – віддаль на яку тягне місток; v – швидкість ковзання На сьогодні актуальна модель Хакслі-Сімонса “ковзаючий філамент-обертаючий місток ”, під кутом 45о – відкритий, під кутом 90о – замкнутий. Неясним залишається лише те, як на молекулярному рівні енергія гідролізу АТФ перетворюється в ковзання протофібрил.
Теорія Давидова (1977) – енергія АТФ (33 кДж·М-1) достатня для збудження внутрішньомолекулярних коливань в молекулі поліпептида Регуляція скелетних м'язів рухом міозину n Міозин зв'язується з відповідним сайтом на актині - це блокується за відсутності Ca2+.
n Періодичне вивільнення Ca2+ дозволяє Tn/TM комплексу обертатися латерально, таким чином, розкривати звязуючий сайт міозину Регуляція скелетних м'язів n Myosin: червоний n Actin: коричневий n Tropomyosin: голубий n Troponin: фіолетовий n Calcium: пісочний n ATP/ADP зелений АКТОМІОЗИНОВА взаємодія Міозин рухається в напрямку кінця зазубрин актинових філаментів Механохімічна енергія йде від гідролізу ATф + ATP, слабо зв'язується ATP висока спорідненість до зв'язування ТМ і ТН модулюють актоміозин
1. Головка міозину зв'язується з відповідною ділянкою актинового філаменту, перша розміщена під кутом 45о 2. Молекула АТФ зв'язується із сайтом на міозині, який тоді дисоціює від актину 3. Міозинова АТФазна гідролізує АТФ, АДФ та Фн знову зв'язуються з міозином 4. Головки міозину пересуваються і зв'язуються з молекулою актину, вони знаходяться під кутом 90о 5. У кінці міозинові головки звільняють АДФ, відновлюючись щільно зв'язуються з ділянкою 6. Звільняється Фн, що ініціює рухову силу. Головки міозину обертаються навколо осі, штовхаючи філаменти актину СЕРЦЕВИЙ М”ЯЗ Міокард складається з скоротливих робочих клітин – кардіоміоцитів, волокон провідної системи і клітин синусно-передсердного та передсердно-шлунучкового вузлів. Кардіоміоцити мають поперечну смугастість, галузяться, відділені один від одного вставними дисками, що свідчить про клітинну будову міокарда. Вставні диски утворені ПМ сусідні клітин, що розділяють міжклітинною щілиною 20 нм. Кардіоміоцит оточений ПМ (7,5 нм) та базальною мембраною (50 нм), разом вони утворюють зовнішню оболонку м'язової клітинисарколему, має Т-систему, міофібрили, СПР, м/х. Значно коротші клітини від м'язових волокон скелетних м'язів; Міокардіоцити розгалужені клітини, які контактують з іншими клітинами контактами, які називають вставними дисками
-електрохімічний імпульс не може поширюватися на всі контактні клітини - відповідає функціональна структура - синцитій (клітини працюють як одна структура) Наявні міофібрили, як і у скелетних м'язах – тому серцевий м'яз володіє смугастістю. Багато кардіоміоцитів є міогенними – імпульс поширюється від клітин м'язів, але не від нервової системи – забезпечує натуральний (природній) ритм серця Електричний зв'язок між клітинами міокарда здійснюється завдяки вставним дискам, що і формує електричний синцитій. Вхідний опір є надзвичайно малим, тому для порогової деполяризації потрібно сильний подразник, крім того збудження охоплює невелику ділянку – захист міокарда. Поширюється збудження на робочий міокард з провідної системи провідними міоцитами (волокнами Пуркін”є).
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 617; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.103.202 (0.009 с.) |