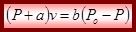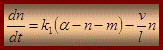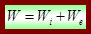Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
ПД поперечносмугастих м”язівСтр 1 из 4Следующая ⇒
ЛЕКЦІЯ №13 на тему: БІОФІЗИКА М”ЯЗІВ ТА КЛІТИННОЇ РУХОМОСТІ Рух – одна з важливих і характерних властивостей живої матерії. Однак біофізичні механізми, що лежать в основі руху є дуже різноманітні. Всі рухові системи поділяють на м'язові та нем'язові. М'язове скорочення – приклад високоспеціалізованого руху, механізми якого добре вивчені на молекулярному рівні. У м'язах основна маса клітинної речовини спеціалізована для виконання скоротливої функції. Два основні білки – актин та міозин становлять близько 80% загального білка м'язів. Актино-міозиновий комплекс – важливий приклад перетворення хімічної енергії у механічну. Розрізняють три типи м'язових систем: m Поперечносмугасті (скелетні) м'язи m Серцевий м'яз; m Гладкі м'язи ПОПЕРЕЧНОСМУГАСТІ М”ЯЗИ Поперечносмугасті скелетні м'язи хребетних тварин складаються з м'язових волокон – багатоядерна клітина циліндричної форми діаметром від 20 до 80 мкм і довжиною декілька міліметрів. Кожне волокно містить близько 200 міофібрил (М) діаметром 1-2 мкм – спеціалізована функціональна ультраструктура поперечносмугастих м'язових волокон. Характерною особливістю будови М є поперечна смугастість, яка зумовлена високою упорядкованістю розміщення тонких та товстих протофібрил. Структура скелетних м'язів БУДОВА М”ЯЗОВОГО ВОЛОКНА Кожна міофібрила за допомогою Z-дисків поділена на структури – саркомери (довжина 2-3-мкм). Товсті та тонкі протофібрили у центрі саркомера утворюють анізотропну смужку (А-диск). Н-зона не містить головок міозину. М-смужка – розташована посередині Н-зони Щодо структурно-функціонального стану, то поперечнопосмуговані м'язові волокна можна поділити на фазні (швидкі та повільні) та тонічні. Фазні м'язові волокна відповідають з поодиноке збудження короткочасним скороченням. Це волокно має видовжену циліндричну форму, до нього можна застосувати кабельну теорію. Rm – 3000-5000 Ом·см2, С – 2,0 мкФ/см2, ПД – -80÷-90 мВ. ПД ПОПЕРЕЧНОСМУГАСТИХ М”ЯЗІВ ПД фазного м'язового волокна має амплітуду 120-130 мВ, овершут – (перевищення над нульовим рівнем МП) – від +30 до +50 мВ, тривалість 3-5 мс. Порогова деполяризація – 15-25 мВ, слідова деполяризація має амплітуду 15-30 мВ, час ранньої слідової деполяризації 10-15 мс (швидкість спаду кінцевої частини слідової деполяризації). Після припинення тетанічного подразнення з'являється значна й тривала пізня слідова деполяризація.
За нормальних умов тонічні м'язові волокна не здатні генерувати ПД, однак після денервації та додавання у зовнішній розчин ТЕА, або іонів Sr2+ або Ba2+ набувають такої здатності до генерації ПД. У відповідь на деполяризацію у тонічних волокнах виникає контрактурне скорочення, яке триває упродовж тривалості деполяризації, це волокно скорочуються в 10 разів повільніше. Зв'язок між збудженням і скороченням у поперечносмугастих м'язах. ПД фазного м'язового волока викликає поодиноке скорочення яке триває понад 300 мс. При збудженні в активації скорочення м'язів головним фактом є деполяризація мембрани, а не іонна природа цього процесу. Деполяризація мембрани викликана ПД поширюється на Т-систему трубочок і стимулює вивільнення іонів Са2+ із саркоплазматичного ретикулуму (СПР). Отже повинна існувати система управління цим процесом, оскільки збудження повинно передаватися вглибину волокна на 10 мкм і синхронізувати роботу усіх міофібрил. А.Хакслі (1959) припустив, що цю роль виконує спеціальна структурно-функціональна система електромеханічного зв'язку: Сарколема; Т-система (має велику кількість інвагінацій ПМ досередини волокна, діаметр цих трубочок 0,03 мкм); СПР; Регуляторні білки тонкої протофібрили Зв'язок між збудженням і скороченням у поперечносмугастих м'язах.
Порожнина Т-трубочок сполучається з зовнішнім середовищем, площа поверхні Т-трубочок у 4 рази більша від площі ПМ. Повноцінний ПД виникає на мембрані Т-системи і здатен збуджувати всі міофібрили волокна в радіальному напрямку. Це зумовлює швидке скорочення волокна за типом “все або нічого”. Деполяризація мембрани Т-трубочок передає і регулює надходження сигналу від ПМ до СПР, завдяки чому вивільняється Са2+ з цистерн останнього. САРКОПЛАЗМАТИЧНИЙ РЕТИКУЛУМ На сьогодні немає задовільного пояснення передачі сигналу від СПР до Т-системи для активації вивільнення Са2+. МОДЕЛІ
Œ Механічна – зміщення негативних зарядів у мембрані Т-трубочок під час деполяризації зсувають містки у тріадах, що спонукає відкривання Са2+ -каналів СПР; Електрична – заряди які проходять через містки, деполяризують мембрани ретикулума, що стимулює відкривання Са2+ -каналів СПР; Ž Хімічна – передача сигналу від Т-системи до СПР здійснюється іонами Са2+ м'язового волокна, або ІТФ. МЕХАНІКА СКОРОЧЕННЯ Ізометричний – напруження активного м'язу не супроводжується скороченням; Ізотонічний – м'яз скорочується, до вільного кінця м'язу прикріплюють вантаж і тоді А =mh. Ізотонічне скорочення. При однаковій вихідній довжині м'язу: a) Ступінь і швидкість укорочення є тим більшим, чим менше навантаження; b) Укорочення досягає свого максимуму тим швидше, чим більше навантаження; c) Чим більше є навантаження, тим пізніше після подразнення починається скорочення і тим раніше воно закінчується. Важливою характеристикою ізометричного скорочення є швидкість вкорочення м'язу V: при Р=0 – Vmax. Наприклад, м'яз жаби скорочується зі V=0,2 м/с, м'язи великих тварин 8 м/с.
ЕНЕРГЕТИКА СКОРОЧЕННЯ За допомогою чутливих термопар А. Хілл виміряв малі кількості виділеного тепла (при поодинокому ізометричному скороченні видіялється 12,54 Дж·кг-1).
де b – константа; Ро – максимальне ізометричне напруження м'яза; а – коефіцієнт теплоти вкорочення; V – швидкість скорочення Рівняння Хілла встановлює зв'язок між напругою та швидкість вкорочення м'яза: Хілл встановив важливий факт, що при ізотонічному скороченні теплота вкорочення QВ, є пропорційною величині вкорочення мяза ∆L:
де а – коефіцієнт теплоти вкорочення
Під час ізотонічного скорочення м'язів хімічна енергія гідролізу АТФ перетворюється в механічну й теплову. Швидке виділення спостерігається на початковій стадії активації скорочення – це теплота активації А, вона виділяється в результаті вивільнення іонів Са2+ із СПР. У міру скорочення мяза виділяється основна кількість теплоти, яка називається теплотою вкорочення QВ, невелика кількість теплоти виділяється чи поглинається при розслабленні м'яза QР, крім цього виконується механічна робота W. ТОВСТІ ПРОТОФІБРИЛИ Основним білкомє міозин – механоензим,що циклічно взаємодіє з актином тонкої протофібрили, який складається з 6 поліпептидних ланцюгів – 2 важких (340 кДа) і 4 легких (140 кДа) ланцюгів мероміозину, С1 та С2 субфрагментів, які входять до складу Важкого мероміозину, до краю Легкого мероміозину протофібрили приєднуються головки, які стирчать з кроком спіралі 43 нм.
МІОЗИН ТА АТФ-азна РЕАКЦІЯ Для виконання механічної роботи м'яз використовує енергію АТФ. Міозинову АТФазу відкрили Енгельгардт й Любимова (1939), кінетика цього ферменту не описується рівнянням Міхаеліса-Ментен. Цей фермент має певні особливості – здатні утворювати інтермедіати (проміжні конформації М* і М** багаті енергією). При взаємодії А-М з АТФ утворюється комплекс АМ**ФАДФ, якщо А заблокований, то М*+АТФ↔М**ФАДФ; Стадія АМ**ФАДФ↔АМ*ФАДФ значно швидша ніж ТЕОРІЯ М”ЯЗОВОГО СКОРОЧЕННЯ Першою була кінетична теорія Хакслі (модель взаємного ковзання двох типів протофібрил, 1954) – товсті та тонкі протофібрили ковзають одна відносно одної, не міняючи довжини, сила яку розвиває саркомер пропорційна кількості мостиків в зоні перекриття фібрил.
Модель Дещеревського (1968) – на товстих протофібрилах є мостики, які замикаються на тонких протофібрилах. Існує три стани мостика: вільний; замкнутий, що тягне з силою f і замкнутий, що гальмує, з силою f. Виходячи з циклічної роботи мостика Дещеревський приходить до рівняння Хілла. ТЕОРІЯ М”ЯЗОВОГО СКОРОЧЕННЯ
де α – загальна кількість мостиків; n – тягнучі мостики; m – гальмуючі мостики; k1 – швидкість переходу з вільного стану до замкнуто тягучих; k2 - швидкість переходу з замкнуто тягучого стану до замкнуто гальмівного; l – віддаль на яку тягне місток; v – швидкість ковзання На сьогодні актуальна модель Хакслі-Сімонса “ковзаючий філамент-обертаючий місток ”, під кутом 45о – відкритий, під кутом 90о – замкнутий. Неясним залишається лише те, як на молекулярному рівні енергія гідролізу АТФ перетворюється в ковзання протофібрил. Теорія Давидова (1977) – енергія АТФ (33 кДж·М-1) достатня для збудження внутрішньомолекулярних коливань в молекулі поліпептида Регуляція скелетних м'язів рухом міозину n Міозин зв'язується з відповідним сайтом на актині - це блокується за відсутності Ca2+.
n Періодичне вивільнення Ca2+ дозволяє Tn/TM комплексу обертатися латерально, таким чином, розкривати звязуючий сайт міозину Регуляція скелетних м'язів n Myosin: червоний n Actin: коричневий n Tropomyosin: голубий n Troponin: фіолетовий n Calcium: пісочний n ATP/ADP зелений АКТОМІОЗИНОВА взаємодія Міозин рухається в напрямку кінця зазубрин актинових філаментів Механохімічна енергія йде від гідролізу ATф + ATP, слабо зв'язується ATP висока спорідненість до зв'язування ТМ і ТН модулюють актоміозин
1. Головка міозину зв'язується з відповідною ділянкою актинового філаменту, перша розміщена під кутом 45о 2. Молекула АТФ зв'язується із сайтом на міозині, який тоді дисоціює від актину 3. Міозинова АТФазна гідролізує АТФ, АДФ та Фн знову зв'язуються з міозином 4. Головки міозину пересуваються і зв'язуються з молекулою актину, вони знаходяться під кутом 90о 5. У кінці міозинові головки звільняють АДФ, відновлюючись щільно зв'язуються з ділянкою 6. Звільняється Фн, що ініціює рухову силу. Головки міозину обертаються навколо осі, штовхаючи філаменти актину
СЕРЦЕВИЙ М”ЯЗ Міокард складається з скоротливих робочих клітин – кардіоміоцитів, волокон провідної системи і клітин синусно-передсердного та передсердно-шлунучкового вузлів. Кардіоміоцити мають поперечну смугастість, галузяться, відділені один від одного вставними дисками, що свідчить про клітинну будову міокарда. Вставні диски утворені ПМ сусідні клітин, що розділяють міжклітинною щілиною 20 нм. Кардіоміоцит оточений ПМ (7,5 нм) та базальною мембраною (50 нм), разом вони утворюють зовнішню оболонку м'язової клітинисарколему, має Т-систему, міофібрили, СПР, м/х. Значно коротші клітини від м'язових волокон скелетних м'язів; Міокардіоцити розгалужені клітини, які контактують з іншими клітинами контактами, які називають вставними дисками -електрохімічний імпульс не може поширюватися на всі контактні клітини - відповідає функціональна структура - синцитій (клітини працюють як одна структура) Наявні міофібрили, як і у скелетних м'язах – тому серцевий м'яз володіє смугастістю. Багато кардіоміоцитів є міогенними – імпульс поширюється від клітин м'язів, але не від нервової системи – забезпечує натуральний (природній) ритм серця Електричний зв'язок між клітинами міокарда здійснюється завдяки вставним дискам, що і формує електричний синцитій. Вхідний опір є надзвичайно малим, тому для порогової деполяризації потрібно сильний подразник, крім того збудження охоплює невелику ділянку – захист міокарда. Поширюється збудження на робочий міокард з провідної системи провідними міоцитами (волокнами Пуркін”є). АВТОМАТІЯ СЕРЦЕВОГО М”ЯЗУ Автоматія – здатність клітин спонтанно деполяризуватися до порогу генерації ПД, це характерно для водіїв ритму. Збудження, що виникає у синусно-передсердному вузлі поширюється в праве передсердя, по спеціальному пучку у ліве передсердя і водночас по міжвузловому тракті до передсердно-шлуночкового вузла, дальше переходить до провідної системи, через яку поширюється на міоцити скоротливого міокарда шлуночків. Швидкість поширення збудження у синусно-передсердному вузлі – 0,1-0,2 м/с; в шлуночках – 0,5-1,0 м/с. МЕХАНІКА СКОРОЧЕННЯ Параметри ізометричного та ізотонічного скорочення відрізняються від характеристик скелетного м'язу. Летальний період ізометричного скорочення в 7-10 разів довший. Швидкість наростання і спаду напруження в передсердях в 2-3 рази більша ніж у шлуночках. Різними сучасними методами показано, що при ізометричному скороченні відбувається внутрішнє вкорочення м'язу (в 3-6 разів більше на відміну від скелетних). ЕНЕРГЕТИКА СКОРОЧЕННЯ При збільшенні вантажу Р, зростає робота W і потужність N м'язу, які досягають максимуму за таких умов Р=0,4 – 0,5 Ро. За подальшого збільшення дані параметри зменшуються, а за Р = Ро вони спадають до нуля. Максимально корисна робота:
Коефіцієнт корисної дії η = 10% при умовах in vitro;
η = 20-30% при фізичному навантаженні; η = 45% у скелетному м'язі.
ГЛАДКІ М”ЯЗИ Гладкі мязи – входять до складу внутрішніх органів і своїм скороченням забезпечують моторику цих органів (травний канал, сечостатева система, кровоносні судини тощо). Основною структурною одиницею гладеньком”язової тканини є міоцит (веретеноподібна м'язова клітина). Довжина міоциту до 20-500 мкм, діаметр 5-10 мкм. q Вісцеральні (гладенькі) м'язи - у внутрішніх органах і судинах - не посмуговані q Міофібрили гладких м'язів – веретеноподібна форма - 20-500 μm in length. q Співвідношення Актину до Міозину ~ 6: 1 у скелетних м'язах, 4: 1 міокардіоцитах ~ 16.5: 1 в гладких м'язах; q Володіють високою еластичністю і здатністю розтягуватися, і надалі здатністю скорочуватися; q Регуляція: гормони, нейротрансмітери, електричним імпульсом, рецепторами. Зовні міоцит вкритий плазматичною мембраною, а на електронно-мікроскопічному зрізі видно своєрідні вигинання – кавеоли, колбоподібні структури й за допомогою вузької частини шийки з'єднуються з зовнішнім міжклітинним середовищем (на 1 мкм2 припадає 30-35 кавеол). Функціональна роль кавеол досі не з'ясована. У деяких ділянках ПМ сусідніх міоцитів є дуже зближеними, ці ділянки нексуси – здійснюють функціональний зв'язок між клітинами, щілина між 2 клітинами становить 2-3 нм. Основними елементами міоциту є ядро, мікротрубочки, лізосоми, СПР, протофібрили.
Потенціал спокою – становить від -50 до -70 мВ; як і в інших м'язових та нервових клітинах в утворенні потенціалу спокою, головним чином, беруть участь іони Na+, K+, Cl-. Усі гладенькі м'язи можуть генерувати потенціали дії: 1) Повноцінні ПД по типу “все або нічого” (фазні); 2) Локальну деполяризацію ПМ (тонічні). ПД можуть бути як простими (тривалістю 20-50 мс, міометрій, ворітна вена, кишківник), так і з вираженими плато (1 с, сечовід) Пороговою концентрацією іонів Са2+ необхідною для активації скорочення гладеньких м'язів є 10-7 М, максимальне скорочення розрізняють при 10-5 М. Товсті протофібрили гладеньких м'язів відрізняються за розміщенням молекул міозину від скелетних. Головним компонентом тонких протофібрил є актин F, котрий полімеризуєтьсяз G-актину, однак в цій протофібрилі немає тропоніну. СКОРОЧЕННЯ q Проста одиниця тканини гладких м'язівє поєднана у синцитій (syncytium) – автономна нервова система іннервує поодинокі прості клітини повністю по всій поверхні або пучками - ПД поширюється через щілинні з'єднання до суміжних клітин q Гладкі м'язи можуть скорочуватися самовільно - це властивість - міогенно ДЖГУТИК Джгутик складається з наступних частин n тіло джгутика - ниткоподібний філамент або флагела; n гак; n базальне тіло. Встановлено, що філамент та гак знаходяться зовні клітинної оболонки, а базальне тіло з’єднує їх з клітиною. В середньому довжина джгутика становить від 10 до 100 мкм з діаметром 23 нм. Нормальні джгутики хвилеподібно вигнуті. Важливим параметром такого джгутика є довжина хвилі філаменту l (l = 2,3 мкм). Існують бактерії з аномальними джгутиками ("кучеряві" джгутики, з l в 2 рази меншою, ніж у нормальних джгутиків, і прямі джгутики). Типи хвилястості джгутиків: а — прямий мутант; б — звивистий мутант; в — нормальний джгутик. Філамент джгутика побудований з ідентичних субодиниць білка – флагеліна. Мономерний флагелін мол. маса 51,5 кДа, асиметрична форма (довжина близько 10 нм). Методами диференціальної мікрокалориметрії і обмеженого протеолізу виявлена доменна будова флагеліну, в якому містяться два периферійні домени Д0 і Д1, що мають 25% a-спіралей, і два центральні домени Д2 і Д3, в яких переважає b-структура. Хімічна структура флагеліну специфічна для кожного штаму бактерій.
Гак служить зв'язуюючою ланкою між базальним тілом і філаментом. Дослідження хімічної структури гаків та базальних тіл показали, що гак побудований з однакових субодиниць, які містять тільки один поліпептид, від 30-42 000 Да. Гак – це структура, що знаходиться у основі філамента, зазвичай вигнутий під кутом і зв’язує філамент з базальним тілом. Зовнішній діаметр гака дещо перевищує діаметр філаменту, загальна довжина якого біля 900 нм. Функція гака ще не цілком зрозуміла, хоча вважають, що він відіграє роль гнучкого з'єднання яке забезпечує ефективну передачу руху від базального тіла до філаменту і водночас надає структурі філамента стійкість по відношенню до змін оточуючого середовища. Стрижень, що виходить з проксимального кінця гака проходить крізь клітинні оболонки і з’єднується з базальними структурами. Базальне тіло - складна структура, яка відповідає за генерацію руху, складається з 9 поліпептидів (9000 до 60000 Да). Ультратонка структура базального тіла грамнегативної бактерії E.coli характеризується наявністю 4 кілець, які контактують з різними структурними елементами мембрани бактерії. Кільце М – прикріплене до ПМ і з нього виходить стрижень джгутика. Кільце S розташовано над ПМ і не з’єднане з жодними іншими клітинними структурами. Кільця L та Р занурені у відповідні шари клітинної стінки і фіксують стрижень, що проходить крізь бактеріальну стінку. Можна припусти, що кільця S та М відповідають за генерацію руху. Механізм роботи джгутика Берг та Андерсен у 1973 році постулювали, що рух бактеріального джгутика є обертовим. Субструктура флагелярного мотора утворюється з багаточисельних копій білків, які синтезуються відповідними генами клітини. Флагелярний мотор – це обертальний мотор, в якому роль “ротора” виконують М- і S- кільця (М-кільце розміщене в плазматичній мембрані), а роль “статора” – трансмембранні білки MotA i MotB, які в плазматичній мембрані оточують М-кільце. М- і S- кільця утворюються переважно з білка FliF (25 копій) і деякої кількості білка FliG. З кільцями М і S жорстко зв’язані стрижень (“ведучий вал”), гак і довгий флагелярний філамент. Кільця L (знаходиться в зовнішній мембрані) і Р (вмонтоване в пептидоглікановий шар) виконують роль “втулки”, в якій з малим тертям і з великою частотою (до 300 Гц) може обертатися стрижень. Тільки S-кільце не контактує з мембранними структурами і знаходиться в міжмембранному просторі. Експериментальні дані вказують на те, що тільки з кільцем М пов'язане обертання джгутика. Нем”язові форми рухомості Мікротрубочки Особливі внутрішньоклітинні структури, будова яких стала відома завдяки застосуванню методів електронної мікроскопії. У 1946 р. Джакус та Холл продемонстрували наявність однакових по діаметру трубчастих структур у війках Раrаmесium. Мікротрубочки це довгі порожнисті циліндри, зовнішній діаметр - 24 нм, а внутрішній – 15 нм. У більшості клітин довжина мікротрубочок зазвичай не перевищує кілька мікрон, хоча в спеціалізованих клітинах, наприклад, у моторних нейронах центральної нервової системи трубочки можуть бути довжиною у кілька сантиметрів. Стінка мікротрубочок (5 нм) побудована з повздовжньо орієнтованих протофібрил, які складаються з глобулярних субодиниць білка тубуліну. Окрім того, що мікротрубочки мають пряме відношення до клітинної рухливості, вони приймають участь і в інших процесах: • в підтриманні форми клітини, • у внутрішньо клітинному транспорті речовин, • в секреції клітинних продуктів, • в рухливості хромосом при поділі клітини і • можливо у реалізації сенсорних зв’язків, • а також в переміщенні компонентів клітинної мембрани. Вони можуть бути розсіяні по всій цитоплазмі або зібрані у організовані структури. Війки та джгутики еукаріот - це спеціалізовані структури, що виходять за межі клітинної поверхні і здатні рухатись. Джгутики та війки можна досить легко відділити від клітин іноді достатньо енергійного струшування клітинної суспензії, що полегшує дослідження їх структури та хімічного складу. Будова аксонеми В центрі знаходиться пара мікротрубочок. Стінка кожної з центральних мікротрубочок утворена 13 повздовжніми рядами білкових субодиниць – протофібрил. Центральну структуру оточують 9 розташованих по кільцю пар мікротрубочок. У дуплеті субфібрила (А) має замкнуту стінку і складається з 13 протофібрил. Субфібрила (В) має серповидний розріз і сполучається з субфібрилою А, складається з 10 протофібрил. Кожна міктротрубочка А з’єднана з мікро-трубочкою В сусіднього дуплета з допомогою білка нексину. Від мікротрубочки А відходить у напрямку до центру радіальна спиця. Додамо, що кожна мікртотрубочка А має подвійний ряд боковий ручок (їх довжина біля 14 нм) побудованих з білка дінеїну і спрямованих до сусіднього дуплету. Всі ці вирости розміщені вздовж субфібрили А на певній відстані: дінеїнові ручки - 24 нм, радіальні спиці - 29 нм і нексинові містки - 86 нм. Спеціалізована структура – базальне тіло – з якої виходить джгутик знаходиться в цитоплазмі клітини біля основи джгутика. В базальному тілі поряд з мікротрубочками А та В є ще мікротрубочки С (10 протофібрилами) так, що по периферії війки розташовуються триплети мікротрубочок. Ці триплети пов’язані між собою боковими виступами і прикріплені до центрального кільця радіальними спицями. Базальне тіло війки чи джгутика по своїй структурі нагадує центріоль. І базальне тіло і центріоль діють як організатори асоціації міктротрубочок. Є ще одна цікава частина джгутика – комірець чи кільцевий валик, що оточує основу джгутика, де він виходить з клітини. На поперечних зрізах комірця в цій ділянці видно волокнисті структури з’єднання між периферичними дуплетами джгутика і внутрішніми шарами клітинної мембрани. Вони жорстко прикріплюють мікротрубочки в клітинній мембрані і беруть участь у русі джгутика Молекулярна структура мікротрубочок Мікротрубочки складаються із білку тубуліну, у вигляді димерів з молекулярною масою 100 кДа. Димери тубуліну розпадаються на мономерні глобули a- і b-тубуліну з М = 50 кДа (450 амінокислотних залишків). Мономери тубуліну полімеризуються, утворюючи довгі протофібрили. Полімерізація починається з утворення димерів тубуліну, а потім b-тубулін одного димера зв'язується з a-тубуліном слідуючого димера. Утворюються стрічки із 13 тубулінових протофібрил, які потім скручуються в замкнуті циліндри, як це маємо в субфібрилі А. Незамкнуті мікротрубочки (субфібрила В) вміщують 10 протофібрил. Нем”язові форми рухомості Амебоїдний рух Амебоїдний рух – характерний для багатьох клітин, таких як амеби та фібробласти. Цей тип руху охоплює всі форми переміщення клітин за допомогою тимчасово утворених на поверхні клітин виростів які мають назву псевдоподій (у фібробластів – ламелоподії). У разі амебоїдного руху основна роль у генерації сили належить актиновим філаментам цитоскелету, при якому відбувається структурні зміни цитоплазми, відомі як переходи гель↔золь. Генерація амебоїдного руху Амебоїдний рух генерується скороченням кортикального шару, що обумовлено функціонуванням мікрофіламентів ектоплазми. Молекулярний механізм такого руху включає декілька етапів: n полімерізацію актину і утворення мікрофіламентів, n прошивання актинових філаментів n утворення гелю. При взаємодії з міозином в присутності АТФ та іонів Са2+ відбувається ковзання актинових філаментів, прикріплених до мембрани, в результаті чого кортикальний шар скорочується.
Після скорочення відбувається деполімеризація актинових філаментів, і цикл повторюється знову. Мікрофіламенти в цитоплазмі амеби є динамічними структурами. Переходи гель «золь в цитоплазмі обумовлені полімеризацією і деполімеризацією актинових філаментів.
ЛЕКЦІЯ №13 на тему: БІОФІЗИКА М”ЯЗІВ ТА КЛІТИННОЇ РУХОМОСТІ Рух – одна з важливих і характерних властивостей живої матерії. Однак біофізичні механізми, що лежать в основі руху є дуже різноманітні. Всі рухові системи поділяють на м'язові та нем'язові. М'язове скорочення – приклад високоспеціалізованого руху, механізми якого добре вивчені на молекулярному рівні. У м'язах основна маса клітинної речовини спеціалізована для виконання скоротливої функції. Два основні білки – актин та міозин становлять близько 80% загального білка м'язів. Актино-міозиновий комплекс – важливий приклад перетворення хімічної енергії у механічну. Розрізняють три типи м'язових систем: m Поперечносмугасті (скелетні) м'язи m Серцевий м'яз; m Гладкі м'язи ПОПЕРЕЧНОСМУГАСТІ М”ЯЗИ Поперечносмугасті скелетні м'язи хребетних тварин складаються з м'язових волокон – багатоядерна клітина циліндричної форми діаметром від 20 до 80 мкм і довжиною декілька міліметрів. Кожне волокно містить близько 200 міофібрил (М) діаметром 1-2 мкм – спеціалізована функціональна ультраструктура поперечносмугастих м'язових волокон. Характерною особливістю будови М є поперечна смугастість, яка зумовлена високою упорядкованістю розміщення тонких та товстих протофібрил. Структура скелетних м'язів БУДОВА М”ЯЗОВОГО ВОЛОКНА Кожна міофібрила за допомогою Z-дисків поділена на структури – саркомери (довжина 2-3-мкм). Товсті та тонкі протофібрили у центрі саркомера утворюють анізотропну смужку (А-диск). Н-зона не містить головок міозину. М-смужка – розташована посередині Н-зони Щодо структурно-функціонального стану, то поперечнопосмуговані м'язові волокна можна поділити на фазні (швидкі та повільні) та тонічні. Фазні м'язові волокна відповідають з поодиноке збудження короткочасним скороченням. Це волокно має видовжену циліндричну форму, до нього можна застосувати кабельну теорію. Rm – 3000-5000 Ом·см2, С – 2,0 мкФ/см2, ПД – -80÷-90 мВ. ПД ПОПЕРЕЧНОСМУГАСТИХ М”ЯЗІВ ПД фазного м'язового волокна має амплітуду 120-130 мВ, овершут – (перевищення над нульовим рівнем МП) – від +30 до +50 мВ, тривалість 3-5 мс. Порогова деполяризація – 15-25 мВ, слідова деполяризація має амплітуду 15-30 мВ, час ранньої слідової деполяризації 10-15 мс (швидкість спаду кінцевої частини слідової деполяризації). Після припинення тетанічного подразнення з'являється значна й тривала пізня слідова деполяризація. За нормальних умов тонічні м'язові волокна не здатні генерувати ПД, однак після денервації та додавання у зовнішній розчин ТЕА, або іонів Sr2+ або Ba2+ набувають такої здатності до генерації ПД. У відповідь на деполяризацію у тонічних волокнах виникає контрактурне скорочення, яке триває упродовж тривалості деполяризації, це волокно скорочуються в 10 разів повільніше.
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 611; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.197.123 (0.153 с.) |