
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекція 8. Видільна системи хребетних тварин (2 год.)
Логіка викладу. 1. Особливості будови видільної систем у анамній. 2. Будова видільної систем у амніот. 3. Ембріональне покоління нирок. Рекомендована література: 1, 2, 3, 4, 5. Видільна система Видільна система виконує дві функції: 1 - стабілізація внутрішнього водно-сольового середовища організму, 2 -виведення метаболітів з організму. Доведено також, що в нирках синтезуються біологічно-активні речовини. Зокрема ренін, який входить до системи ренін-ангіотензиноген. Для життєдіяльності тканин тіла необхідне відповідне середовище, найбільш важливою особливістю якого є наявність в оточуючій клітини рідині певної кількості й у певному співвідношенні деяких солей в розчиненому вигляді. Іони цих солей легко проникають через мембрану клітини, тому і в крові, і в тканинній рідині їх співвідношення завжди однакове. Підтримка рівноваги між тим, що потрапило в організм (через травну систему чи шкіру) і тим, що видаляється - вкрай важлива функція та умова існування живого. Цю функцію виконують сольові залози зябрового апарата, ректальна залоза акул, носові або очні залози водних птахів та черепах, потові залози ссавців, ниркові канальці. Видільна система, крім видалення води й солей, виводить метаболіти - шкідливі продукти обміну речовин, перш за все продукти білкового обміну, зокрема високотоксичні іони амонію. У печінці амоній перетворюється в менш токсичну сечовину та сечову кислоти. Сечовина - продукт азотистого трубка з частиною целомічної порожнини автономізується, а ще пізніше зв'язок трубки з целомом поривається. Видільна система ланцетника дещо нагадує протонефридійну систему плоских червів (мал.1).
Мал. 1. Видільна система ланцетника: 1 - каналець з декількома воронками і "полум'яними"клітинами — соленоцитами; II- "полум 'яні "клітини - солено-цити (велике збільшення).
У ланцетника є клітини - соленоцити, які з'єднуються з виносною трубочкою, що відкривається в атріальну порожнину тіла. Соленоцити збирають метаболіти з целомічних мішків, які знаходяться над зябровими щілинами. Продукти розпаду потрапляють в атріальну порожнину і з потоком води виводяться назовні. У порожнині тіла поряд із нефридіями кровоносні судини утворюють густу сітку капілярів у вигляді клубочка. Отже, видільні органи ланцетника нагадують метанефридії кільчастих червів й одночасно мають риси подібності з нефронами хребетних. З підвищенням рівня обміну речовин в еволюційному ряду хребетних вдосконалювались і органи виділення. Видільна система хребетних тварин утворена трьома основними типами нирок: переднирка, первинна і вторинна нирки (мал.2).
і
А - пронефроса з зовнішнім клубочком; Б - мезонефроса з внутрішнім клубочком і В - метанефроса ссавців. 1 - зовнішній клубочок; 2 - внутрішній клубочок; 3 - нефростом; 4 — звивистий каналець; 5 - петля Генле; 6 — збірний каналець.
І хоча часто їх розглядають як окремі самостійні утвори, насправді між ними існує ряд проміжних форм. Згідно з концепцією про те, що нирки хребетних слід розглядати як безперервне розмаїття морфологичних і функціональних одиниць, можна почати опис спочатку гіпотетичного типу нирки, яку називають голонефросом. Суть цього полягає в тому, що в усіх тулубових сегментах мезодерма має у своєму складі з кожного боку проміжний відділ, представлений в кожному сегменті окремим нефростомом – невеликим утвором, що з'єднує соміт і бічну пластинку. Очевидно, у первісної форми хребетних кожен нефростом давав початок одному нирковому сегменту, включаючи до його складу невелику частину пророжнини целому. Ця стінка целому ставала шаром капсули Боумена, а від останньої відходив звичайний каналець. Отже, вздовж тіла з кожного боку тулуба знаходився ряд метамерних ниркових елементів. З обох боків утворились два протоки, які збирали сечу від ряду сегментарних елементів і ззаду об'єднувались перед відкриттям назовні тіла або в клоаку. Ці канали назвали архінефритичними, вольфовими або пронефритичними. Найбільш примітивним типом нирки є пронефрос. Така нирка добре розвинена й функціонує в зародків риб і амфібій. У зародків птахів і ссавців цей орган являє собою декілька пар рудиментарних метамерних канальців або двох клітинних тяжів. Так, пронефрос амфібій - це 3-5 пар канальців, що відповідають рівням 3-4 сомітів. Типовий каналець пронефроса виникає з нефростому, який відділяється від соміту, має форму стебельця з внутрішньою порожниною, якою сполучається з целомічною порожниною. Латеральні кінці кількох канальців пронефроса об'єднуються й утворюють зачаток первісного ниркового протоку, який далі проростає в бік клоаки. Пронефрос знаходиться поблизу головного кінця тіла й дещо нагадує голонефрос личинок міксин та безногих амфібій. Видільна система дорослих міксин ще слабо відрізняється від голонефросу, але й у них найперші канальці редуковані. В усіх вищих риб та амфібій пронефрос дегенерується і його функцію бере на себе мезонефрос або тулубова нирка - новий орган, який розміщений дещо каудальніше. У амніот позаду мезонефросу розвивається третій тип видільної системи (органа) -метанефрос або тазова нирка чи справжня нирка. Кожна з цих структур є парним органом, який розміщуєтеся позаперитонеально (згідно з походженням!). У птахів та ссавців у процесі ембріогенезу послідовно з'являються й зникають і пронефрос, і мезонефрос, що відбиває основні етапи розвитку хордових. У ембріонів птахів та ссавців функціонує мезонефрос (тулубова нирка), але подібно пронефросу дегенерує за винятком його протоків.
Розглянемо спочатку розвиток нирок в анамній. В ембріонів круглоротих, подібно до інших хребетних, закладаються й функціонують головні нирки (pronephros). Пізніше позаду від них розвиваються парні тулубові нирки (mesonephros), що функціонують у дорослих особин. Вони лежать на спинному боці порожнини тіла у вигляді стрічкоподібних утворів під статевою залозою (мал. 3).
Мал. 3. Поздовжній розріз задньої частини тулуба міноги: 1- мезонефрос; 2 - сечопровід; 3 - сечостатевий синус; 4 - статева пора; 5 - сечостатевий отвір;6 - кишечник; 7 - анальний отвір; 8 - статева залоза.
По нижньому краю кожної нирки проходять сечоводи -вольфові канали, які ведуть у сечостатевий синус, що відкривається на сечостатевому сосочку сечостатевим отвором. У більшості круглоротих пронефрос редукується: від нього залишається кілька канальців, які відкриваються в навколосерцеву сумку. У бделостом зберігається частина переднирки і в тулубовому відділі. Нирки круглоротих мікроанатомічно суттєво відрізняються від нирок щелепноротих, являючи собою нібито першу фазу виникнення гломерулярного апарату (мальпігієвих тілець). Упродовж усієї нирки проходить у вигляді шнура гломус, що являє собою слабо впорядковану сукупність артеріальних капілярів, які виділяють фільтрат. Останній по міжклітинних проміжках стікає в короткі ниркові канальці, де відбувається часткова реабсорбція корисних для організму речовин з фільтрату. Отже, у круглоротих ще не відбулось мікроанатомічного об'єднання обох елементів фільтруючого клубочка та капсули, що приймає фільтрат. У щелепноротих будова нирки набагато складніша. У ній, перш за все, значно зростає кількість канальців (набагато більше, ніж сомітів); по-друге, у самців частина вивідного каналу нирки функціонує як вивідний протік для сперми. Нирка анамній звичайно видовженої форми і тягнеться майже вздовж більшої частини тіла або позаду від залягання пронефросу. Але навіть у примітивних форм акул і химер передній її кінець тонкий і основний об'єм нефрогенної тканини знаходиться ззаду. Причина, очевидно, полягає в тому, що сім'яник у них встановлює зв'язок з передньою частиною нирки, використовуючи канальці мезонефросу для виведення сперми, і ця частина вже не функціонує як нирка. Це, по суті, нагадує придаток сім'яника ссавців. У самців ця частина нирки дегенерує, а задня розростається і втрачає метамерність. Між клубочками та пронефричним каналом тепер з'являються збірні (вивідні) трубочки, в які виливається сеча з декількох клубочків.
Особливою є видільна система променевоперих риб. У них сім'яник має свій вивідний канал, який зовсім не зв'язаний із видільною системою, тому нирка має відносно велику довжину (мал.4).
Мал. 4. Вивідні капали сечостатевої системи самця щуки. 1 - нирка; 2 - сечовід; 3 - сечовий міхур; 4 - зовнішній сечовий отвір; 5 — сілі 'яник; 6 - сім япровід; 7 - зовнішній статевий отвір; 8 — кишечник; 9 - анальний отвір.
Кардинальна морфрологічна відміна мезонефросу або тулубової нирки від пронефросу (головної) чи, тим паче, голонефросу (гіпотетичної нирки) хордових полягає в тому, Що канальці в ній безпосередньо зв'язані з кровоносною системою. На вільному кінці мезонефричного каналу утворюється чашоподібне заглиблення — гломерулярна або боуменова капсула, в яку занурений клубочок капілярів (гломерулярне тільце Мальпігі). У пронефросу це був васкуляризований валик (гломус). У зародків і передня частина мезонефросу не функціонує як типова нирка, а в дорослих форм він редукується. Саме в процесі диференціації задньої частини мезонефросу передня частина дегенерує. Сама задня частина нефрогенної тканини для утворення мезонефросу не використовується, а створює несегментовану сферичної форми масу у верхній стінці поперекового відділу порожнини тіла. У цьому скупченні мезодерми починає диференціюватись велика кількість ниркових канальців, із яких пізніше утворюється тазова нирка або метанефрос. Ці канальці вже не впадають в архінефричний канал, бо для них утворюється новий канал (уретра). Він у філогенезі з'являється пізно, а в ембріогенезі закладається раніше, ніж канальці, і, як гадають, індукує їх появу. Він бере початок з того місця, де канал пронефросу впадає в клоаку, росте краніально і вростає в метанефрос та продовжує в ньому багаторазово поділятись (мультиплікує) на дрібні, так звані збираючі трубочки. Далі до верхівок цих трубочок приєднуються канальці нефрону.
Нефрон – це функціонально-структурна одиниця нирок. Він складається з тільця Мальпігі та відносно довгої трубочки. Судинний клубочок утворений приносним артеріальним капіляром у боуменовій капсулі. У ссавців поверхневий шар нирки (кіркова речовина) усього кілька міліметрів завтовшки, вона охоплює внутрішню частину -мозкову речовину. У мозковій речовині розрізняють окремі ділянки трикутної форми, які радіально покреслені. Це так звані ниркові піраміди (мал. 5).
А -розріз через нирку людини; Б- схема циркуляції крові в нирці. 1 - ниркові піраміди; 2 - розміщення нефрона; З - ниркова миска; 4 - сечовід; 5 - ниркова артерія; 6 — ниркова вена; 7 - капсула Боумена; 8 - мальпі-гісв клубочок; 9 - петля Генле; 10 - збірний каналець.
їх верхівки обернені до ниркової лоханки, а основи - до кіркової речовини. Пірамідки віддалені одна від одної кірковою речовиною, яка заходить сюди від поверхневого шару, утворюючи ниркові стовпи. Ниркова артерія входить у ворота нирки, розгалужується на гілки, які проходять між нирковими пірамідами до їх основи. Їх називають частковими артеріями. Дійшовши до основи пірамід, часткові артерії поділяються на дві гілки, які розходяться, ідуть вздовж основи двох сусідніх пірамід приблизно на межі між ними та кірковою речовиною. Вони названі дуговими артеріями. Від цих артерій у напрямі до поверхні нирки відходять численні гілочки - часткові артерії, і від них ідуть короткі бічні гілочки, які несуть кров до мальпігієвих клубочків. Їх і називають приносними артеріями, які переходять в капіляри, що утворюють судинний клубочок Мальпігі (чудові сітки). Далі капіляри збираються в одну артерійку, котра називається виносною. Остання вужча за приносну й саме тому в клубочку виникають умови для підвищеного кров'яного тиску, котрий сприяє фільтрації рідини з крові в капсулу Боумена. Капсула - закритий з усіх боків мішечок, що оточує мальпігіїв клубочок. Та частина мішечка, яка прилягає до капілярів клубочка, або внутрішня її стінка досить тоненька і складається із зірчастих клітин - подоцитів, між відростками яких мають місце вузенькі щілини, так що місцями від порожнини між шарами капсули капіляри відділені лише базальною мембраною. Мальпігієві тільця бувають лише в корковому шарі нирки. Порожнина капсули переходить у просвіт звивистого канальця, в якому розрізняють проксимальний і дистальний відділи. Ці звивисті канальці густо обплетені сіткою капілярів, які щойно покинули клубочок і несуть, по суті, артеріальну кров або залишки фільтрату. Фільтрат клубочка відрізняється від крові відсутністю в ньому форменних елементів великих молекул білка. Підраховано, що на протязі доби утворюється у безхвостої амфібії близько півлітра, а в людини біля 200 літрів первинної сечі - фільтрату клубочка. Але в нирках відбувається не лише процес фільтрації, а й процес реабсорбції (зворотного всмоктування) завдяки ферментативним реакціям. Тому вторинна сеча у тієї ж амфібії складає 5 % від первинної, а в людини - біля 1 %. При цьому сильно змінюється її склад.
Глюкоза повністю повертається в кров, повністю змінюється кількісний склад інших елементів. Клубочки та канальці усіх хребетних можна поділити на три розряди: 1 - клубочки відносно великі, кількість фільтрату, що виділяється ними, теж велика (більшість кісткових риб, пластинозяброві риби, амфібії); 2 - клубочки малі або зовсім відсутні, а в деяких костистих риб відсутній також дистальний звивистий канадець. Фільтрату виділяється мало (морські костисті риби та рептилії); 3 - клубочки відносно великі, але між звивистими канальцями першого та другого порядків з'являється петля Генле. Первинної сечі виділяється багато, а вторинної - мало (ссавці, слабше виражено це у птахів). Звичайно, дані варіанти нефронів пов'язані з життям у середовищах з різною концентрацією солей або з можливою втратою води через шкіру. Прісноводні риби й амфібії живуть у середовищі з нижчою концентрацією солей, ніж у тканинах тіла, тому вода має тенденцію проникати в організм, а великі клубочки здатні виділити з організму зайву воду. Окрім того їх зябра здатні всмоктувати солі з води. У морських риб і міног концентрація солей у зовнішньому середовищі набагато перевищує їх концентрацію у внутрішньому середовищі. Частково морські кісткові риби, латимерія та міноги солі та деякі продукти азотистого обміну виділяють через зябра. Економія води в їх організмі здійснюється за рахунок зменшення розміру ниркових тілець або їх редукції. Акули, як і прісноводні риби, воду виводять великими нирковими клубочками. Солі вони виводять сольовою ректальною залозою. Крім того, акули здатні зберігати в крові велику кількість кінцевих продуктів азостистого обміну у вигляді сечовини. Виняток, який поки що залишається без пояснення, складають міксини. Концентрація солей в рідинах їх тіла така ж, як і в оточуючому морському середовищі. Наземні тварини живуть у сухому середовищі, вода постійно втрачається з поверхні тіла, що загрожує зневодненням і підвищенням концентрації солей у тканинах тіла цих тварин. Океанічні рептилії і птахи п'ють морську воду й тому мають залози для виведення солей з організму (птахи -носові залози, а черепахи - очні). Ссавці здатні економити воду інакше. Вони мають великі ниркові тільця і в канальці спочатку поступає багато рідини - фільтрату, але далі там відбувається поверенння води. У ссавців для цього з'являється петля Генле, яка опущена в мозковий шар нирки. Тут фізіологи відкрили явище протитічності - в петлі Генле в дистальному звивистому канальці йде процес реабсорбції. У птахів також є, хоч і слабо розвинена, петля Генле, але в них, як і в рептилій, кінцеві продукти азотистого обміну виділяються у вигляді сечової кислоти, яка майже нерозчинна у воді, і тому для їх транспорту не потрібно багато води. У людини біля 1 млн. ниркових клубочків, у птахів та хатньої миші - близько 200 тис. Збираючі трубочки в нирці ссавців виносять сечу в ниркові чашечки, які далі передають її в ниркову лоханку. У риб, амфібій, рептилій і частково в птахів є ворітна система нирок. Клубочок утворюється артеріальною системою, а не венозною (в артеріальній крові більший тиск, що важливо для фільтрації). У багатьох хребетних є сечовий міхур. У риб це порожнина для накопичення сечі перед виведенням її назовні тіла. В акул і примітивних костистих риб такі утвори невеликі, слабо примітні й утворюються з вивідних протоків. У кісткових риб (дводишних і нижчих променевоперих) невеликий сечовий міхур утворений з архінефричних каналів. У тетрапод сечовий міхур - гігієнічний утвір. У безхвостих амфібій, можливо черепах, зворотне відсмоктування води з міхура попереджає обсихання тіла. Це вивороти клоаки, які не мають зв'язків з сечоводами. Сечовий міхур відсутній у деяких ящірок, змій та крокодилів і птахів (крім австралійського страуса ему). У ссавців сечовий міхур тісно пов'язаний з видільною системою. Сечоводи впадають безпосередньо в сечовий міхур - сильно розтягнутий мішок, всередині вистелений перехідним епітелієм. Еволюція видільної системи хребетних відбувається поступово шляхом ембріонального відщеплення від уже існуючих канальців нових, досконаліших. Комплекс початкових, порівняно просто побудованих канальців становить переднирку й зберігається тільки в передній частині тіла зародків. Він розвинувся раніше від інших ниркових канальців і тому відокремився й спеціалізувався як зародковий видільний орган. У сегментах, розміщених позаду, початкові канальці редукувалися, а комплекс додаткових канальців подальших генерацій являє собою первинну нирку. Ці канальці первинної нирки побудовані складніше і вже мають, окрім секреторного апарату, ще й фільтраційний. Нефростоми в більшості канальців зникають. Подальшим ускладненням є те, що вивідна протока почала давати від себе бічні вирости та нові генерації канальців, які вступають у зв'язок з цими виростами. Задній виріст набуває у вищих хребетних переважного значення (сечовід). Він на кінці розгалужується і туди впадає велика кількість вторинних ниркових канальців, кожен з яких починається мальпігієвим тільцем. Ця частина органа відокремлюється і стає вторинною ниркою, яка розвивається повільніше й починає функціонувати пізно, тому первинна нирка спеціалізується як зародковий орган виділення. Отже, еволюція видільної системи йде в напрямку збільшення кількості нефронів, диференціації канальців на відділи, поєднання секреторної та фільтруючої частини й відокремлення видільної системи від статевої.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 887; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.36.30 (0.037 с.) |
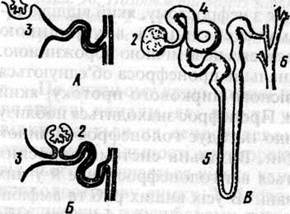 Мал. 2. Схема ниркових канальців:
Мал. 2. Схема ниркових канальців:

 Мал. 5. Нирка ссавця (за Смітом).
Мал. 5. Нирка ссавця (за Смітом).


