Для осуществления вдоха и выдоха необходимо преодолеть сопротивление дыханию. Различают эластическое и неэластическое сопротивление.
Эластическое сопротивление значительно больше неэластического. 2/3 его величины создается поверхностным натяжением жидкости, выстилающей альвеолы, 1/3 – эластическими компонентами органов грудной и брюшной полостей. Эластическое сопротивление больше на вдохе, чем на выдохе. Измеряется тем градиентом давления (Δ Р), который нужно приложить, чтобы заполнить легкие определённым объёмом воздуха (Δ V):
R эл = Δ Р / Δ V
Неэластическое сопротивление формируют:
1. Аэродинамическое сопротивление (Rа) – обусловлено трением слоев воздуха о стенки воздухоносных путей и друг о друга. Rа составляет 80-90% величины неэластического сопротивления.
Rа зависит от длины (l), просвета (r) дыхательных путей и плотности воздуха (η), что описывается уравнением Пуазейля:
R а = 8 l η / π r4
Неэластическое сопротивление больше на выдохе, чем на вдохе. Это является одной из причин большей продолжительности выдоха по сравнению с вдохом (в среднем в 1,3 раза).
Аэродинамическое сопротивление при дыхании через нос, который играет роль клапана, более чем на 50% превосходит его величину при ротовом дыхании. В последнем случае Rа равно приблизительно 1-2 см вод. ст. × c × л -1.
2. Вязкое сопротивление тканей грудной и брюшной полостей – определяется их внутренним трением и неупругой деформацией. Составляет 10-20% величины неэластического сопротивления.
3. Инерционное сопротивление – его величина очень мала.
В соответствии с законом Хагена-Пуазейля неэластическое сопротивление измеряется тем градиентом давления (ΔР), который необходим для того, чтобы придать потоку воздуха определённую объёмную скорость (Q):
R неэл = Δ Р / Q
При прочих равных условиях сопротивление дыханию зависит от его паттерна (pattern, англ. – образец, модель), под которым понимают различные характеристики дыхательного цикла: длительность фаз, глубина дыхания, скорость изменения давления и перемещения воздуха в дыхательных путях. Так, при глубоком дыхании увеличивается доля эластического сопротивления, а при частом – неэластического.
В ходе преодоления сопротивления дыхательные мышцы совершают определенную работу (А):
А = Р × V,
где Р – создающееся давление (в альвеолах),
V – объём лёгких во время вдоха.
Если при совершении работы давление изменяется, то необходимо применять интеграл ∫PdV.
В покое дыхательные мышцы совершают работу, равную 0,1-0,5 кг×м/мин, и при этом потребляют 2-2,5% поглощенного кислорода. При нагрузке выполняемая работа возрастает до 1-5 кг×м/мин. В этих условиях на сокращение дыхательных мышц затрачивается около 20% потреблённого кислорода (до 30-40% при затруднённом дыхании). Это ущемляет потребность других органов в кислороде. Поэтому существует «кислородный потолок» – то количество кислорода, которое организм может затратить на работу дыхательной мускулатуры. В связи с этим имеется и «предел дыхания» – объём вентиляции легких, увеличение сверх которого энергетически невыгодно, т.к. весь дополнительно потребляемый кислород будет расходоваться на обеспечение работы дыхательных мышц. Равен 100-120 л/мин.
Зарисуйте спирограмму, опишите лёгочные объёмы и емкости, укажите их величину. Минутная вентиляция лёгких. Альвеолярная вентиляция. Какой из этих параметров наиболее адекватно отражает эффективность внешнего дыхания и почему?
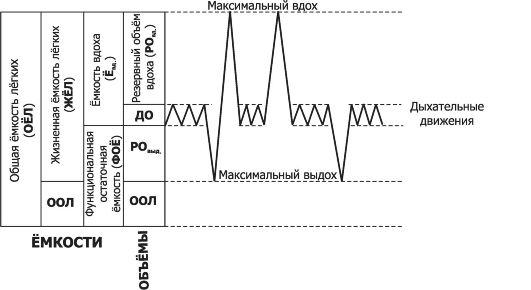
К статическим относят лёгочные объёмы:
1. Дыхательный объём (ДО) – количество воздуха, вентилируемое при спокойном дыхании. У взрослого человека он составляет приблизительно 0,5 л. Экспираторный объём несколько меньше инспираторного, поскольку кислорода поглощается больше, чем выделяется углекислого газа.
2. Резервный объём вдоха (РОВд) – количество воздуха, которое можно вдохнуть дополнительно после спокойного вдоха (около 3 л).
3. Резервный объём выдоха (РОВыд) – количество воздуха, которое можно выдохнуть дополнительно после спокойного выдоха (около 1,3 л).
4. Остаточный объём (ОО) – количество воздуха, остающееся в легких после максимально глубокого выдоха (около 1,2 л). Даже при открытом пневмотораксе после выхождения большей части ОО, называемой коллапсным объёмом, в спавшихся лёгких остаётся некоторое количество воздуха – минимальный объём. Этот воздух находится в «воздушных ловушках», образующихся в результате того, что часть бронхиол спадается раньше альвеол. Это используется в судебной экспертизе для решения вопроса о том, дышал ли ребенок после рождения. Если да, то его лёгкие не тонут в воде.
Из лёгочных объёмов формируются ёмкости:
1. Жизненная ёмкость лёгких (ЖЕЛ) – наибольшее количество воздуха, которое можно выдохнуть после максимально глубокого вдоха
ЖЕЛ = ДО + РОВ д + РОВ ыд
Является показателем подвижности лёгких и грудной клетки. Зависит от:
– возраста: в процессе онтогенеза уменьшается, особенно после 40 лет;
– пола: у женщин на 25% меньше, чем у мужчин;
– роста: ЖЕЛ(л) = k × рост (м) (k для мужчин – 2,5, для женщин – 2,0);
– положения тела: в вертикальном несколько больше, чем в горизонтальном;
– степени тренированности: у спортсменов выше, чем у нетренированных людей.
Должная величина ЖЕЛ может быть рассчитана по формуле Людвига:
для мужчин:
ЖЕЛ = 40 × рост (см) + 30 × масса (кг) – 4400;
для женщин:
ЖЕЛ = 40 × рост (см) + 10 × масса (кг) – 3800.
2. Резервная ёмкость вдоха (РЕВд) – максимальное количество воздуха, которое можно вдохнуть после обычного выдоха
РЕВ д = ДО + РОВ д
3. Функциональная остаточная ёмкость (ФОЕ) – количество воздуха, остающееся в лёгких после спокойного выдоха
ФОЕ = РОВ ыд + ОО
ФОЕ имеет важное физиологическое значение, поскольку выравнивает колебания содержания О2 и СО2 в альвеолярной смеси газов, которые могли бы быть вызваны сменой фаз дыхательного цикла. Это происходит следующим образом. При вдохе к альвеолам поступает дыхательный объем за вычетом объема анатомического мертвого пространства. К последнему относят воздухоносные пути, в которых не происходит газообмен, – от полости носа до 22 поколения дихотомического деления бронхов. Его объем равен 100-200 мл, в среднем 2 мл на кг массы тела, т.е. 150 мл при массе 75 кг. Эта порция (500-150=350 мл) смешивается с воздухом, содержащимся в легких, количество которого в среднем у молодых людей составляет 2,4 л, у пожилых 3,4 л. Поэтому при вдохе обновляется лишь 1/7 часть смеси газов в альвеолах. В результате состав последней существенно не изменяется. Следовательно, ФОЕ обеспечивает постоянство состава альвеолярной газовой смеси, что необходимо для эффективного протекания газообмена в легких.
4. Общая ёмкость лёгких – количество воздуха, которое содержится в них на высоте максимального вдоха. Складывается из ЖЕЛ и ОО. У взрослого человека составляет 4,2-6,0 л.
Лёгочные объёмы, за исключением ОО, как и ЖЕЛ и РЕВд, могут быть измерены с помощью спирометрии и пневмотахометрии. В последнем случае регистрируют не сами объемы, а объемную скорость струи воздуха.
ФОЕ можно измерить только непрямыми методами – разведения гелия, вымывания азота, интегральной плетизмографии.
К динамическим показателям внешнего дыхания относятся:
1. Лёгочная вентиляция (ЛВ) – характеризуется минутным объемом дыхания (МОД), т.е. количеством воздуха, вентилируемым за 1 мин. МОД зависит от ДО и частоты дыхания (ЧД), которая у взрослого человека в среднем составляет 14 в 1 мин (от 10 до 18), у новорождённого – 40-60 в 1 мин.
МОД = ДО × ЧД
При ДО = 500 мл и ЧД = 14 в мин МОД равен 7 л. При физической нагрузке он может увеличиваться до 120 л
В связи с тем, что к альвеолам, как уже отмечалось, поступает не весь ДО, МОД не может характеризовать эффективность дыхания в целом. Для этого необходимо оценить альвеолярную вентиляцию.
2. Альвеолярная вентиляци я (АВ) – часть МОД, достигающая альвеол. Отличается от ЛВ на величину вентиляции мёртвого пространства (ВМП):
АВ = ЛВ – ВМП
При ЧД = 14 в мин:
АВ = (500 - 150) × 14 ≈ 5 л в мин
При частом и поверхностном дыхании, несмотря на достаточную величину ЛВ, АВ будет снижена, т.к. в этом случае будет вентилироваться, главным образом, «мёртвое» пространство. Поскольку объём последнего постоянен, то АВ тем лучше, чем глубже дыхание. Следовательно, эффективность внешнего дыхания (ЭВД) зависит от соотношения АВ и МОД.
ЭВД = АВ / МОД × 100
В покое ЭВД составляет 60-70%.
3. Максимальная вентиляция лёгких (МВЛ) – объем воздуха, проходящий через легкие за определенный промежуток времени (обычно за 10 сек) при дыхании с максимальной частотой (40-60 в мин) и глубиной. МВЛ зависит от возраста, пола, конституции. При пересчете на минуту у молодых людей МВЛ составляет 120-170 л.
4. Максимальные скорости вдоха и выдоха – измеряются путём пневмотахометрии или пневмотахографии.
5. Отношение объёма форсированного выдоха за 1 сек к ЖЕЛ (проба Тиффно) – у молодых людей 70-80%, с возрастом снижается до 65-70%.
6. Бронхиальное сопротивление – определяется, в частности, с помощью форсированной ЖЕЛ (ФЖЕЛ), которая изучается в условиях максимально быстрого выдоха. В норме разница между ЖЕЛ и ФЖЕЛ составляет 100-300 мл. При нарушении бронхиальной проходимости она увеличивается.
7. Работа дыхательных мышц.
8. рН и бикарбонаты крови – их уровень связан с напряжением углекислого газа.
9. Парциальное давление кислорода и углекислого газа в альвеолярной смеси газов и их напряжение в артериальной крови.
Рецепторы лёгких, дыхательных путей и дыхательных мышц. Их физиологическое значение. Рефлекс Геринга-Брейера. Теоретические основы различных видов искусственного дыхания и закрытого массажа сердца при оказании неотложной помощи.
Различают постоянные и непостоянные (эпизодические) рефлекторные влияния на функциональное состояние дыхательного центра.
Постоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Геринга — Брейера), корня легкого и плевры (пульмоторакальный рефлекс), хеморецепторов дуги аорты и сонных синусов (рефлекс Гейманса), проприорецепторов дыхательных мышц.
Наиболее важным рефлексом является рефлекс Геринга — Брейера. В альвеолах легких заложены механорецепторы растяжения и спадения, являющиеся чувствительными нервными окончаниями блуждающего нерва. Любое увеличение объема легочных альвеол возбуждает эти рецепторы.
Рефлекс Геринга — Брейера является одним из механизмов саморегуляции дыхательного процесса, обеспечивая смену актов вдоха и выдоха. При растяжении альвеол во время вдоха нервные импульсы от рецепторов растяжения по блуждающему нерву идут к экспираторным нейронам, которые, возбуждаясь, тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает условия для повышения возбудимости инспираторной части дыхательного центра и осуществлению активного вдоха. Значение рефлексов Геринга-Брейера заключается в регуляции соотношения глубины и частоты дыхания в зависимости от состояния легких. Это повышает экономичность работы дыхательной системы. Рефлексы Геринга-Брейера имеют важное значение у новорожденных (в первые 3-4 дня). У взрослого человека они реализуются только при дыхательных объемах свыше 1 л. Кроме того, они обеспечивают тахипноэ при гиперкапнии и гипоксии, поскольку после «выключения» блуждающих нервов, например, анестетиками, в этих условиях развивается лишь гиперпноэ.
Кроме того, активность инспираторных нейронов усиливается при нарастании концентрации углекислого газа в крови, что также способствует проявлению вдоха.
Пульмоторакальный рефлекс возникает при возбуждении рецепторов, заложенных в легочной ткани и плевре. Проявляется этот рефлекс при растяжении легких и плевры. Рефлекторная дуга замыкается на уровне шейных и грудных сегментов спинного мозга.
К дыхательному центру постоянно поступают нервные импульсы от проприорецепторов дыхательных мышц. Во время вдоха происходит возбуждение проприорецепторов дыхательных мышц и нервные импульсы от них поступают в инспираторную часть дыхательного центра. Под влиянием нервных импульсов тормозится активность вдыхательных нейронов, что способствует наступлению выдоха.
Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждениемразнообразных экстеро- и интерорецепторов. К ним относятся рефлексы, возникающие при раздражении рецепторов слизистой оболочки верхних дыхательных путей, слизистой носа, носоглотки, температурных и болевых рецепторов кожи, проприорецепторов скелетных мышц. Так, например, при внезапном вдыхании паров аммиака, хлора, сернистого ангидрида, табачного дыма и некоторых других веществ происходит раздражение рецепторов слизистой оболочки носа, глотки, гортани, что приводит к рефлекторному спазму голосовой щели, а иногда даже мускулатуры бронхов и рефлекторной задержке дыхания.
При раздражении эпителия дыхательных путей накопившейся пылью, слизью, а также попавшими химическими раздражителями и инородными телами наблюдается чиханье и кашель. Чиханье возникает при раздражении рецепторов слизистой оболочки носа, кашель — при возбуждении рецепторов гортани, трахеи, бронхов.
При внезапном прекращении сердечной деятельности и дыхания наступает
состояние клинической смерти. Это может случиться при поражении
электротоком, утоплении и в ряде других случаев при сдавливании или
закупорке дыхательных путей. Однако если сразу же приступить к непрямому массажу сердца и искусственной вентиляции легких, то в ряде случаев удается спасти пострадавшего. Наиболее эффективно использование для искусственной вентиляции легких специальных аппаратов, с помощью которых в легкие вдувается воздух. При отсутствии таких аппаратов искусственную вентиляцию легких проводят различными способами, из которых наиболее распространен способ «изо рта в рот».
Прежде чем начать эту процедуру, надо уложить пострадавшего на спину,
чтобы его воздухоносные пути были свободны для прохождения воздуха. Для этого его голову максимально запрокинуть назад. Для этого подкладывают одну руку под шею, а другой надавливают на темя. В результате корень языка отодвигается от задней стенки гортани и восстанавливается проходимость дыхательных путей.
При сжатых челюстях надо выдвинуть нижнюю вперед и, надавливая на
подбородок, раскрыть рот, затем очистить салфеткой ротовую полость от слюны или рвотных масс и приступить к искусственной вентиляции: на открытый рот пораженного положить в один слой салфетку (носовой платок), зажать ему нос, сделать глубокий вдох, плотно прижать свои губы к губам пораженного, создав
герметичность, с силой вдуть воздух ему в рот. Вдувать надо такую
порцию воздуха, чтобы она каждый раз вызывала возможно более полное
расправление легких, что обнаруживается по движению грудной клетки. Небольшие порции воздуха не дадут никакого эффекта. Воздух вдувают ритмично 16—18 раз в минуту до восстановления естественного
дыхания. Эффективный массаж сердца можно проводить только в том случае, если больной лежит на жесткой поверхности. При проведении массажа на улице больного укладывают спиной на асфальт или жесткие носилки, при проведении массажа в помещении — на пол или длинный стол. Если остановка сердца произошла у больного, находящегося в мягкой кровати, опускают изголовье (убирают подушки), под спину подкладывают поднос, лист фанеры и т. п. Массаж сердца может быть успешным только при одновременном проведении искусственного дыхания.
Техника. Непрямой массаж сердца, проводимый одним человеком.
Пострадавшего укладывают на спину, запрокидывают ему голову. Платком или марлей очищают полость рта, делают 3—5 дыханий методом рот в рот, рот в нос. Располагаются слева от больного или опускаются на колени, если он лежит на полу (на земле). На нижнюю треть грудины кладут ладонь левой руки, а сверху нее — ладонь правой руки. Надавливают на грудину достаточно сильно, чтобы она опустилась приблизительно на 5 см, за тем задерживают на мгновение в этом положении и быстро отпускают. Цикл надавливание—отпускание должен составлять несколько меньше секунды. Через каждые 15 циклов делают 2 частых дыхания методом рот в рот или рот в нос. Признаком эффекта массажа является сужение ранее расширенных зрачков, появление фибрилляции желудочков, а затем и редких сердечных сокращений, восстановление спонтанного дыхания. Массаж продолжают до полного восстановления сердечной деятельности, появления отчетливых сердцебиений и пульса на периферических артериях.
14.Опишите, в чем заключается газообмен в лёгких. Напишите формулу Фика. Исходя из данной формулы, перечислите, какие факторы способствуют газообмену в легких, а какие препятствуют. Укажите парциальное давление газов в альвеолярном воздухе и их напряжение в артериальной и венозной крови капилляров.
Мы вдыхаем атмосферный воздух, содержащий около 21% кислорода, 0,03% углекислого газа, остальное — азот и небольшое количество инертных газов и водяного пара. В составе выдыхаемого воздуха кислорода около 16%, углекислого газа — около 4%. Итак, в легких богатый кислородом атмосферу, поступившее во время вдоха, заменяется на воздух, в котором содержание кислорода в 1,3 раза меньше, а содержание углекислого газа больше аж в 133 раза. Организм человека в состоянии покоя ежеминутно получает 250-300 мл кислорода и выделяет 250-300 мл углекислого газа.
За сутки диффундирует около 500 л кислорода и 430 л углекислого газа. Количественные закономерности этого процесса выражаются первым законом диффузии Фика, сформулированным им для диффузии через клеточные мембраны в 1885 г:
M / t = ΔP / (x/SK α),
где М – количество газа,
t – время,
M/ t – объёмная скорость диффузии, равная 15-30 мл/мин в покое, 60-70 мл/мин при нагрузке,
Δ P – разница парциального давления газа в двух точках,
х – расстояние между этими точками,
S – площадь газообмена,
К – коэффициент диффузии,
α – коэффициент растворимости газа.
Следовательно, скорость диффузии прямо пропорциональна ΔP – силе, обеспечивающей направленное движение молекул газа, и обратно пропорциональна x/SKα – величине сопротивления диффузии.
Применительно к условиям газообмена в легких ΔP есть разница между парциальным давлением газа в альвеолярной газовой смеси и его напряжением в крови легочных капилляров.
Парциальным давлением (ПД) называется то давление, которое оказывал бы данный газ, если бы он один занимал весь объем смеси газов.
По закону Дальтона оно пропорционально процентному содержанию газа в смеси и общему давлению смеси. ПД не зависит от природы газа. Состав альвеолярной газовой смеси отличается от состава вдыхаемого и выдыхаемого воздуха тем, что в нем меньше всего кислорода и больше углекислого газа.
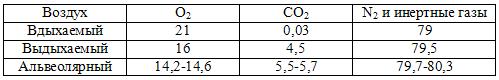
Напряжением газа в жидкости называется сила, с которой молекулы растворенного газа стремятся выйти в газовую среду.
Парциальное давление и напряжение О2 и СО2 в лёгких

15.Вентиляционно-перфузионные соотношения в различных отделах легких и их значение. Понятие об анатомическом и физиологическом мертвом пространстве. Особенности кровообращения в легких.
Эффективность выполнения основной функции дыхательной системы зависит от соответствия перфузии (кровотока) в регионах легких с их вентиляцией. Так, хороший кровоток будет недостаточным для газообмена в регионах легких, если они слабо вентилируются воздухом с низким содержанием кислорода.
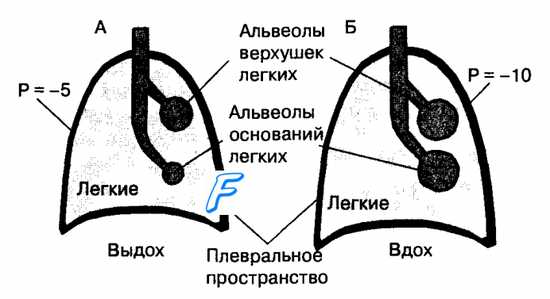
У человека в вертикальном положении тела величина легочного кровотока на единицу объема ткани легкого линейно убывает в направлении снизу вверх, и меньше всего снабжаются кровью верхушки легких. Соответственно в положении тела человека на спине кровоток в нижних (дорсальных) отделах легких становится выше, чем в верхних (вентральных). Это обусловлено тем, что артериальная кровь, поступающая в легкие из правого желудочка, проходит по сосудам легких из областей низкого внутриплеврального давления в области тонкостенных капилляров, которые окружены альвеолами, содержащими воздух под давлением, близким к атмосферному. Поэтому в зависимости от соотношения давления в альвеолах (РА), мелких артериях (Ра) и мелких легочных венах (Pv) легкие разделены на функциональные зоны Веста.
В верхушках легких (зона 1) могут возникнуть области с давлением в легочных капиллярах (особенно в фазу диастолы) ниже альвеолярного (Ра > Ра > Pv)- Капилляры в таких зонах могут спадаться, и кровоток через них становится невозможным. Такие участки легких вентилируются, но не участвуют в газообмене и формируют альвеолярное мертвое пространство. В средних отделах легких (зона 2) под действием гравитации давление в альвеолах, как правило, превышает венозное (Ра > РА > Pv). Поэтому величину кровотока в зоне 2 по Весту определяет разность между артериальным и альвеолярным давлениями. В зоне 2 практически не возникает альвеолярное мертвое пространство. В нижних отделах легких (зона 3) давление в легочных венах выше альвеолярного (Ра > Pv > Ра) и величина кровотока, как и в обычных сосудах, определяется разницей между артериальным и венозным давлениями.
Дыхательное мертвое пространство — объем дыхательных путей, в котором не происходят процессы газообмена между воздухом и кровью.
Различают анатомическое и физиологическое (или функциональное) мертвое пространство.
Анатомическое дыхательное мертвое пространство представляет собой объем воздухоносных путей, начиная от отверстий носа и рта и кончая дыхательными бронхиолами легкого.
Под функциональным (физиологическим) мертвым пространством понимают все те участки дыхательной системы, в которых не происходит газообмена. К функциональному мертвому пространству в отличие от анатомического относятся не только воздухоносные пути, но также альвеолы, которые вентилируются, но не перфузируются кровью. В таких альвеолах газообмен невозможен, хотя их вентиляция и происходит.
Существенной особенностью сосудистой системы легких является то, что она включает сосуды малого круга и бронхиальные артерии большого. Первые служат для газообмена, вторые обеспечивают кровоснабжение ткани легких. У человека между ними имеются анастомозы, роль которых в гемодинамике малого круга значительно возрастает при застойных явлениях в нем. Легочная артерия разветвляется на более мелкие артерии, а затем артериолы. Артериолы окружены паренхимой легких, поэтому кровоток в них тесно связан с режимом вентиляции легких. В легких имеется 2 типа капилляров: широкие, диаметром 20-40 мкм, и узкие – 6-12 мкм. Стенка легочного капилляра и альвеолы образуют функциональную единицу – альвеолокапиллярную мембрану. Через нее осуществляется газообмен. Минутный объем крови в сосудах малого круга такой же, как и большом, но кровяное давление меньше. Оно не может значительно повышаться из-за большой растяжимости стенок сосудов легких.
16.Нарисовать кривую диссоциации оксигемоглобина. Описать точки «зарядки» и «разрядки». Каково влияние прямых факторов на сродство гемоглобина к кислороду.
Связывание О2
97% О2, транспортирующегося от легких к тканям, химически связано с гемоглобином.
Молекула гемоглобина состоит из 4 субъединиц, поэтому реакция оксигенации протекает следующим образом: Hb + 4О2 = Hb(О2)4. Молекула О2 обратимо связывается с гемом гемоглобина. Каждый грамм гемоглобина может связать 1,34 мл О2 (число Хюфнера) (1,39 мл, если гемоглобин химически чистый). Следовательно, при содержании гемоглобина в 1 л крови 140 г в этом объеме будет находиться (без учета физически растворённого О2) 1,34×140 = 190 мл О2, т.е. 19 об.%. Количество О2, которое вообще способна связать кровь, называется кислородной ёмкостью крови (в среднем 20 об.%). Реализуется полностью, если кровь контактирует с газовой смесью с высоким содержанием О2 (РО2 > 300 мм рт. ст.). В естественных условиях гемоглобин оксигенирован только на 97%.
Насыщением (saturation) гемоглобина кислородом называется процентное отношение HbО2 к общему содержанию гемоглобина.
SО2 = [HbО2] / [Hb]+[HbО2]
Реакция оксигенации гемоглобина, как указывалось, подчиняется закону действующих масс. Это означает, что отношение между гемоглобином и HbО2 зависит от содержания физически растворенного О2 в крови, которое, в свою очередь, в соответствии с законом Генри-Дальтона, пропорционально напряжению О2. Графически эту зависимость отражает кривая диссоциации оксигемоглобина (КДО).
Еще в 1904 г. С.Bohr установил, что характер реакции гемоглобина с О2 описывается S-образной кривой. Известны следующие причины особой конфигурации кривой.
В 1921 г. G.Adair предположил, что соединение гемоглобина с О2 происходит в 4 этапа, из которых самый медленный – первый (гипотеза «промежуточных соединений»). Поэтому S-образная форма кривой обусловлена кооперативным взаимодействием полипептидных цепей.
Возможны и другие объяснения, согласно которым существуют 2 состояния гемоглобина, переходящие друг в друга при изменении содержания кислорода в результате либо конформационных перестроек, либо присоединения или отщепления низкомолекулярных веществ. Параметры равновесия реакций оксигенации для этих двух форм гемоглобина различны, что и объясняет S-образную форму кривой.
На кривой выделяют 3 участка: от 0 до 10 мм рт. ст. – прямо пропорциональная зависимость, от 10 до 60 – насыщение очень быстрое, от 60 до 90 – насыщение почти не изменяется. В артериальной крови (напряжение О2 96 мм рт. ст.) гемоглобин насыщен кислородом на 97%, в венозной крови (напряжение О2 40 мм рт. ст.) – на 75%.
В настоящее время принято оценивать положение КДО не по наклону кривой, а по расположению на ней двух точек. Первая соответствует 50%-ому насыщению гемоглобина кислородом. Это Р50 – напряжение полунасыщения (точка разрядки по Крогу). В норме при рН = 7,4 и t = 37оС Р50 артериальной крови 26 мм рт. ст. (3,46 кПа). Оно выше у женщин, чем у мужчин. Напряжение разрядки достаточно высоко, что обеспечивает эффективный градиент напряжения О2 между капиллярами и тканями, где напряжение О2 не более 10-15 мм рт.ст. Вторая точка – Р95 – точка зарядки – соответствует 95%-ому насыщению гемоглобина кислородом. В норме Р95 артериальной крови 70 мм рт. ст.
Значение S-образной формы кривой.
1. Крутой наклон среднего участка, соответствующий напряжениям О2 в тканях, благоприятствует отдаче О2 в них.
2. Пологая часть, соответствующая высоким напряжениям О2, обеспечивает стабильность тканевого напряжения О2 в условиях, когда напряжение О2 артериальной крови может уменьшаться: при подъеме в горы или на самолете, при заболеваниях легких, с возрастом. Даже когда парциальное давление О2 в альвеолярной смеси газов снижается до 60 мм рт. ст., гемоглобин в артериальной крови все еще насыщен О2 на 89%, что только на 8% ниже нормальной 97% сатурации.
3. С другой стороны, когда парциальное давление О2 в альвеолярной газовой смеси увеличивается до 500 мм рт. ст. (при вдыхании воздуха под повышенным давлением, например, на глубине моря или в компрессионных камерах), насыщение гемоглобина кислородом также изменяется мало – максимум на 3%.
Следовательно, при варьировании парциального давления О2 в альвеолах от 60 до 500 мм рт. ст. его напряжение в тканях изменяется только на несколько мм рт. ст. Таким образом, гемоглобин буферирует сдвиги тканевого напряжения О2.
КДО может занимать различное положение, характеризующееся смещением ее средней части вправо или влево. Смещение влево сопровождается снижением Р50 и повышением сродства гемоглобина к О2 (СГК), а вправо – увеличением Р50 и уменьшением СГК.
Прямое влияние оказывают химические вещества, которые могут взаимодействовать с гемоглобином и изменять конформацию его молекулы. Это лиганды (О2, Н+, СО2, органические и неорганические ионы).
I. В 1907 C.Bohr, A.Krogh установили зависимость положения КДО от содержания СО2 в крови. Сначала «эффект Бора» связывали только с СО2. Однако затем J.Barcroft (1909) показал, что подобная связь существует и при воздействии других кислот. В настоящее время эффект Бора рассматривают как влияние рН среды на взаимодействие гемоглобина с различными лигандами.
При снижении рН (увеличении [Н+]) СГК падает. Поэтому КДО смещается вправо. Уменьшение оксигенации гемоглобина при снижении рН – эффект Рута.
Описанные явления объясняются тем, что при изменении рН происходит сдвиг константы диссоциации, степени ионизации и конформации групп, связанных с О2 в тетрамере гемоглобина. В результате этого меняется СГК. Различают респираторный и метаболический компоненты эффекта Бора.
Метаболический – связан с изменением рН под влиянием кислых или щелочных продуктов обмена веществ.
Респираторный – связан с влиянием СО2 на КДО. Образующийся при тканевом метаболизме СО2 диффундирует внутрь эритроцитов, где под влиянием карбоангидразы образуется Н2СО3. Появляющиеся при ее диссоциации (Н2СО3 → Н+ + НСО3-) протоны приводят к снижению рН. Это рН-зависимый эффект СО2. Кроме того, СО2 образует карбаминовые связи с концевыми группами α и β цепей гемоглобина, способствуя таким образом стабилизации дезоксиформы гемоглобина. Это рН-независимый эффект СО2.
На выраженность эффекта Бора влияют: 1) органические фосфаты – при повышении содержания 2,3-дифосфоглицерата (2,3-ДФГ) падает коэффициент Бора, который определяют как Δ log Р50 / Δ log рН; 2) температура; 3) насыщение крови О2; 4) электролиты; 5) свойства Hb; 6) пол - коэффициент Бора меньше у мужчин, чем у женщин.
Таким образом, эффект Бора – это результат сложного взаимодействия рН, СО2, 2,3-ДФГ и других указанных факторов.
Биологический смысл эффекта Бора состоит в том, что он способствует поглощению О2 в легких и отдаче его в тканях.
В лёгких – поглощение О2 идет одновременно с выделением СО2, поэтому по мере насыщения гемоглобина кислородом КДО сдвигается влево, т.е. СГК возрастает. В результате скорость диффузии О2 из альвеол в кровь несколько увеличивается.
В тканях – одновременно с выходом О2 в эритроциты поступает СО2, поэтому КДО смещается вправо. Это приводит к снижению СГК и благоприятствует отдаче О2 тканям.
II. В 1967 г. была описана специфическая роль органических фосфатов в эритроцитах. 2,3-ДФГ, значительно уступая АТФ в энергетической ценности, играет огромную роль в регуляции транспорта О2. Во-первых, он является аллостерическим модулятором – взаимодействует с β-цепями восстановленного гемоглобина благодаря соответствию зарядов некоторых аминокислотных остатков последнего распределению зарядов в органическом фосфате. После образования связи с β-цепью 2,3-ДФГ закрывает вход в центральный канал молекулы, что уменьшает СГК. Во-вторых, 2,3-ДФГ изменяет внутриклеточный рН.
На реакцию 2,3-ДФГ и гемоглобина влияют:
1) соотношение Hb/HbО2 – чем оно выше, тем больше связывается 2,3-ДФГ;
2) рН –ж снижение рН увеличивает сродство гемоглобина к 2,3-ДФГ;
3) рСО2 – уменьшение рСО2 повышает сродство гемоглобина к 2,3-ДФГ;
4) температура – ее возрастание препятствует присоединению 2,3-ДФГ к гемоглобину;
5) анионы – могут конкурировать с 2,3-ДФГ за одни и те же центры связывания.
В эритроцитах имеется автономная система регуляции образования 2,3-ДФГ, работающая по принципу обратной связи. С изменением содержания 2,3-ДФГ в эритроцитах связана зависимость СГК от их возраста - молодые эритроциты имеют меньшее СГК, по мере старения оно возрастает, поскольку уровень 2,3-ДФГ уменьшается.
Роль 2,3-ДФГ в качестве регулятора положения КДО заключается не только во влияниии на СГК. Он служит посредником других влияний (рН, рСО2, температуры, ионного состава) на кислородтранспортную функцию крови.
III. Органические и неорганические соли – при увеличении их концентрации КДО сдвигается вправо, поскольку от солевого состава среды зависит конформация макромолекулы гемоглобина.
К косвенным факторам, как уже отмечалось, относят рН, влияние которого на СГК рассмотрено выше, и температуру. Уменьшение температуры увеличивает интенсивность образования HbО2, т.е. при прочих равных условиях смещает КДО влево. Повышение температуры, напротив, снижает СГК, и КДО смещается вправо.
Температурный коэффициент, представляющий собой отношение Δ log рО2 / Δ log t, зависит от рН, содержания 2,3-ДФГ, насыщения крови кислородом. Он различен у мужчин и женщин. Колеблется от 0,018 до 0,024.
17.Нарисовать кривую диссоциации оксигемоглобина. Описать точки «зарядки» и «разрядки». Каково влияние косвенных факторов на сродство гемоглобина к кислороду. Эффект Вериго-Бора и его биологическое значение.
Реакция оксигенации гемоглобина, как указывалось, подчиняется закону действующих масс. Это означает, что отношение между гемоглобином и HbО2зависит от содержания физически растворенного О2 в крови, которое, в свою очередь, в соответствии с законом Генри-Дальтона, пропорционально напряжению О2. Графически эту зависимость отражает кривая диссоциации оксигемоглобина (КДО).
Еще в 1904 г. С.Bohr установил, что характер реакции гемоглобина с О2 описывается S-образной кривой. Известны следующие причины особой конфигурации кривой.
В 1921 г. G.Adair предположил, что соединение гемоглобина с О2 происходит в 4 этапа, из которых самый медленный – первый (гипотеза «промежуточных соединений»). Поэтому S-образная форма кривой обусловлена кооперативным взаимодействием полипептидных цепей.
Возможны и другие объяснения, согласно которым существуют 2 состояния гемоглобина, переходящие друг в друга при изменении содержания кислорода в результате либо конформационных перестроек, либо присоединения или отщепления низкомолекулярных веществ. Параметры равновесия реакций оксигенации для этих двух форм гемоглобина различны, что и объясняет S-образную форму кривой.
На кривой выделяют 3 участка: от 0 до 10 мм рт. ст. – прямо пропорциональная зависимость, от 10 до 60 – насыщение очень быстрое, от 60 до 90 – насыщение почти не изменяется. В артериальной крови (напряжение О2 96 мм рт. ст.) гемоглобин насыщен кислородом на 97%, в венозной крови (напряжение О2 40 мм рт. ст.) – на 75%.
В настоящее время принято оценивать положение КДО не по наклону кри





