Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Теория «скольжения» мышечного сокращения. Кризис теории «скольжения». Теория «вкручивания», объясняющая механизм мышечного сокращения.
Головка миозина в покое представляет собой комплекс «миозин + АДФ + фосфат». Активные центры актина обладают большим сродством к этому комплексу, в результате чего происходит присоединение головки миозина к активным центрам актинового филамента. Присоединение актина вызывает быстрое освобождение АДФ и фосфата из миозина, что приводит к изменению конформации головки. При этом головка поворачивается на 45о вокруг своей оси (рабочий ход). Так как головка имеет несколько центров связывания, то они последовательно взаимодействуют с активными участками на актиновом филаменте и при этом развивается тянущее усилие. После поворота головки к ней вместо ушедших АДФ и фосфата присоединяется АТФ, образуя комплекс «миозин + АТФ». Актин обладает к этому комплексу малым сродством, в результате чего происходит отсоединение головки миозина (разрыв поперечных мостиков). Головка становится перпендикулярно к актиновому филаменту. В головке миозина, уже не связанной с актином, происходит гидролиз АТФ, вновь образуется комплекс «миозин + АДФ + фосфат» и головка вновь способна присоединяться к актиновому филаменту. В каждый конкретный момент развития сокращения часть головок миозинового филамента соединена с активными центрами актинового филамента, а другая часть свободна, что позволяет последовательно осуществлять скольжение актиновых нитей вдоль миозиновых, уменьшение длины саркомеров и укорочение мышцы в целом (или развитие напряжения мышцы). В настоящее время появились некоторые факты, которые не вписываются в классическую теорию скольжения: 1) миозиновый филамент при сокращении меняет свою длину и диаметр (утолщается и укорачивается); 2) при сокращении укорачивается не только диск I, но и диск А; 3) между актиновыми и миозиновыми филаментами расстояние – 13 нм, а длина головки миозина – 19-21 нм; 4) сокращение мышцы ступенчатое, а не плавное, как при скольжении. Эти и другие факты позволили Н.С.Мирошниченко и М.Ф.Шуба (1990) высказать гипотезу, что скольжение актиновых филаментов вдоль миозиновых невозможно из-за структурных препятствий и специфического характера действия сил на сократительный аппарат мышцы. Авторы считают, что в основе сокращения лежит вкручивание миозиновых филаментов в трубкообразные структуры, образованные актиновыми филаментами, а само сокращение обеспечивается междоменными перемещениями в работающих по очереди головках миозина. Указанная гипотеза вкручивания в настоящее время находит всё большее и большее признание у специалистов по «молекулярной механике» и может стать новой современной теорией сокращения скелетных мышц в будущем.
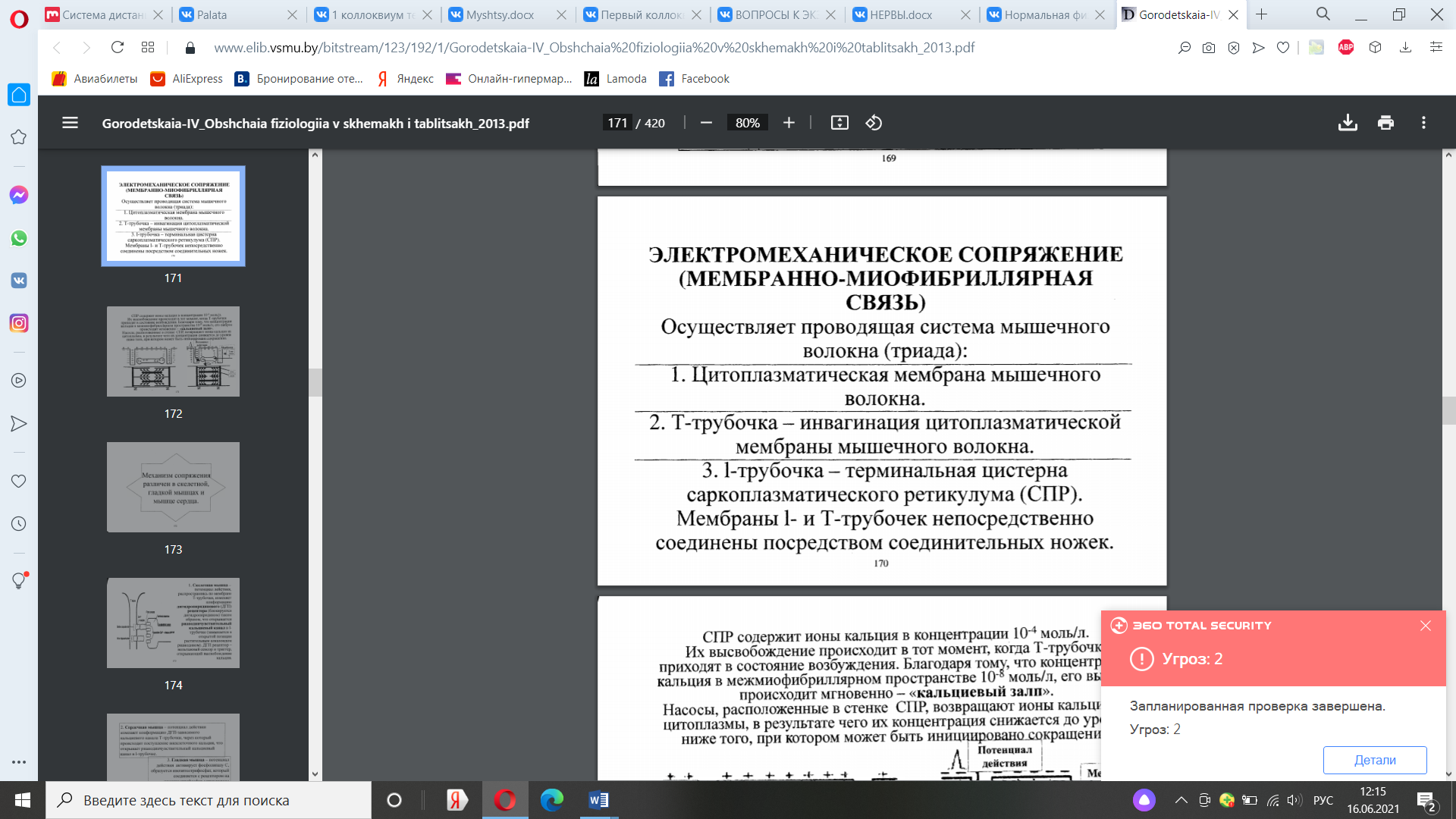
21. Компоненты проводящей системы мышечного волокна (мембранно-миофибриллярная связь). Механизм сопряжения возбуждения и сокращения в мышце. Каковы его особенности в скелетной, гладкой и сердечной мышцах? Роль Са2+ и Н+ в мышечном сокращении.
Механизм сопряжения различен в скелетной, гладкой мышцах и мышце сердца. 1. Скелетная мышца - потенциал действия, распространяясь по мембране Т-трубочки, изменяет конформацию дигидропиридинового (ДГП) рецептора (блокируется дигидропиридином) таким образом, что открывается рианодинчувствительный кальциевый канал в Iтрубочке (замыкается в открытой позиции растительным алкалоидом рианодином). ДГП рецептор - вольтажный сенсор и триггер, открывающий высвобождение кальция. 2. Сердечная мышца - потенциал действия изменяет конформацию ДГП-зависимого кальциевого канала Т-трубочки, через который происходит поступление внеклеточного кальция, что открывает рианодинчувствительный кальциевый канал в 1-трубочке. 3. Гладкая мышца - потенциал действия активирует фосфолипазу С, образуется инозитолтрифосфат, который соединяется с рецептором на инозитолтрифосфат-управляемом (лиганд-зависимом) кальциевом канале СПР, что его открывает. Роль кальция и протонов. Кальций, как уже указывалось, инициирует мышечное сокращение. При гидролизе АТФ появляются ____ протоны. Они конкурируют с кальцием за карбоксилаты тропонина и вытесняют его: ТпССООСа + Н+ -> ТпССООН+Са ++ Кальций откачивается в СПР. Защелка, по-прежнему, остается в / неионизированном, т.е. «включенном» состоянии. Следовательно, протоны поддерживают мышечное сокращение. При ресинтезе АТФ протоны потребляются, защелка переходит в ионизированное состояние и препятствует взаимодействию нитей.
21. Энергообразование и теплообразование в скелетной мышце. Энергия для сокращения мышц образуется при гидролизе АТФ под влиянием фермента АТФ-азы. Свойства этого фермента появляются у головки миозина (тяжёлых цепей), когда она присоединяется к активным центрам актина. В расслабленном состоянии ни миозин, ни актин не обладают АТФ-азной активностью. При расщеплении АТФ меняется конформация АТФ-азного центра головки миозина и головка переходит в новое высокоэнергетическое состояние. Она поворачивается и присоединяется к активному центру актинового филамента. Повторное присоединение головки миозина к актину сопровождается новым поворотом головки. При каждом цикле соединения и разъединения головки миозина с актином гидролизуется одна молекула АТФ на один поперечный мостик. При этом концентрация кальция в миоплазме должна быть не менее 10-5 М. Одна молекула АТФ при расщеплении до АДФ и Н3РО4 выделяет 9,57 ккал (40 кДж) энергии. Распад АТФ, т.е. снижение отношения АТФ к АДФ, запускает ресинтез АТФ (принцип Энгельгарта). Ресинтез АТФ осуществляется анаэробными и аэробными путями. Для этого в мышцах имеется три энергетических системы, которые включаются последовательно по мере расходования АТФ: 1) анаэробная фосфагенная система (АТФ-КФ система); 2) гликолитическая система; 3) окислительная система. Эти системы отличаются друг от друга по энергетической ёмкости, т.е. по максимальному образованию энергии в единицу времени. Как только АТФ гидролизовалась до АДФ, мгновенно (через несколько миллисекунд) включается фосфагенная система и происходит срочный анаэробный ресинтез АТФ из креатинфосфата (КФ), т.к. КФ значительно больше, чем АТФ (креатинкиназная реакция): АДФ + КФ → АТФ + К Если истощаются запасы КФ, то АТФ образуется из двух молекул АДФ (миокиназная реакция): 2АДФ → АТФ + АМФ АМФ подвергается дезаминированию: АМФ → ИМФ + NH3. Одновременно идёт ресинтез КФ в митохондриях. Ёмкость фосфагенной системы мала. При её максимальной работе АТФ хватает на 5-6 секунд работы. Если сокращение мышц продолжается, то последовательно развёртываются гликолитическая и окислительная системы. Работу гликолитической системы запускает АДФ. Анаэробно начинают расщепляться глюкоза и гликоген до лактата. При этом одна молекула глюкозы даёт энергию для синтеза трёх молекул АТФ. Эта АТФ расходуется на работу мембранных насосов. Образование АТФ анаэробным путём происходит в 2-3 раза быстрее, чем аэробным. Ёмкость гликолитической системы в 25 раз больше, чем фосфагенной, но намного меньше, чем окислительной. Поэтому сокращение мышц при анаэробном гликолизе может быть интенсивным, но будет продолжаться 1-2 минуты, а затем с накоплением молочной кислоты наступает утомление. При продолжающемся сокращении мышц через 2-3 минуты развёртывается окислительная система и ресинтез АТФ будет осуществляться в основном за счёт окислительного фосфорилирования. При этом одна молекула глюкозы даёт 38 молекул АТФ (2 молекулы АТФ образуется в результате анаэробного этапа и 36 молекул АТФ – аэробный этап). Ёмкость окислительной системы в тысячи раз превышает ёмкость фосфагенной и гликолитической систем. Поэтому при хорошем кровоснабжении и достаточном поступлении кислорода мышцы работают несколько часов без утомления. Если сокращение мышц длительное, но малоинтенсивное, а потребность мышц в кислороде при этом удовлетворяется полностью, то АТФ ресинтезируется системой окислительного фосфорилирования за счёт 58 окисления жиров. Такая ситуация наблюдается у спортсменов-стайеров (бег на марафонские дистанции). При интенсивном сокращении мышц (выполнение большой работы за короткое время) энергия для сокращения мышц выделяется за счёт окисления углеводов гликолитической системой. Такая ситуация наблюдается во время бега на короткие дистанции у спортсменов-спринтеров.
|
||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 513; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.146.255.127 (0.007 с.) |