Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Акустическая рефлексометрия
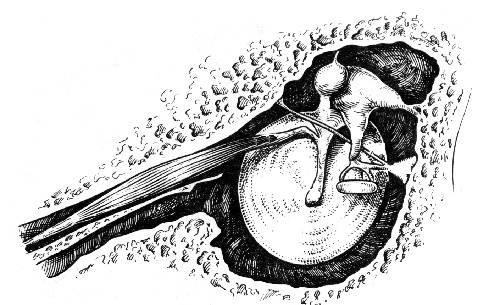
Среднее ухо содержит две мышцы, прикрепляющиеся к цепи слуховых косточек — мышцу, напрягающую барабанную перепонку и стременную. Стременная мышца (M. stapedius) — самая маленькая в организме: ее средняя длина — 6,3 мм, а площадь поперечного сечения — 4,9 мм2. Мышца полностью заключена в пирамидальный выступ (eminentia pyramidalis) на задней стенке барабанной полости и берет начало от стенок своего собственного канала. Ее сухожилие выходит через верхушку пирамидального выступа и далее идет горизонтально до прикрепления к задней поверхности шейки стремени. Таким образом, при своем сокращении стременная мышца тянет стремя назад. Стременная мышца иннервируется стременной ветвью VII (лицевого) черепного нерва, отходящей от него непосредственно вблизи самой мышцы. Мышца, напрягающая барабанную перепонку (M. tensor tympani) располагается в костном полуканале непосредственно над слуховой трубой, от которой она отделена тонкой костной перегородкой. Мышца начинается от хряща слуховой трубы, от стенок собственного канала, части основной кости, прилегающей к стенке канала. При своем выходе из канала сухожилие этой мышцы делает поворот вокруг маленького крючкообразного выступа на промонториуме (processus cochleariformis), пересекает барабанную полость в латеральном направлении и прикрепляется к вершине рукоятки или шейке молоточка. При сокращении мышцы, напрягающей барабанную перепонку, рукоятка молоточка смещается кпереди и внутрь. Эта мышца иннервируется одноименной веткой мандибулярного нерва (3-я ветвь V – тройничного – черепного нерва), волокна которой проходят через ganglion oticum. Обе внутрибарабанные мышцы полностью заключены в костные каналы и связаны с цепью слуховых косточек соответствующими сухожилиями. Это, по мнению Г. фон Бекеши, позволяет избежать искажений звука, возникающих при мышечных сокращениях. При гистологическом исследовании мышц среднего уха найдены как поперечнополосатые, так и гладкие волокна. Что не удивительно, поскольку указанные мышцы, функционирующие при звуковом раздражении непроизвольно, у некоторых людей могут сокращаться по их желанию.
Известно, что некоторые мышцы сокращаются в ответ на звук. Рефлекторное сокращение мышц ушной раковины, например, называется pinna reflex. Прэйер (Preyer, 1879) подробно изучал этот феномен у грызунов. Гораздо большее значение имеют сокращения мышц барабанной полости в ответ на звуковые раздражения. Впервые двустороннее сокращение мышцы, напрягающей барабанную перепонку, при воздействии на одно ухо звуком большой интенсивности было обнаружено в конце 19 века в ходе экспериментов на собаках. Позже (в начале 20-го века) было показано, что стременная мышца кошек и кроликов также отвечает на интенсивную звуковую стимуляцию.
Первым сокращения стременной мышцы у человека наблюдал через перфорированную барабанную перепонку Люшер (Luscher) в 1929 г. Изучением акустического рефлекса также занимались Политцер, Кобрак, Линдсей, Лоренте де Но, Филип, Холлпайк и другие известные ученые. Сокращение мышц барабанной полости в ответ на интенсивную звуковую стимуляцию получило название акустического (мышечного) рефлекса - АР. Он носит безусловный характер.
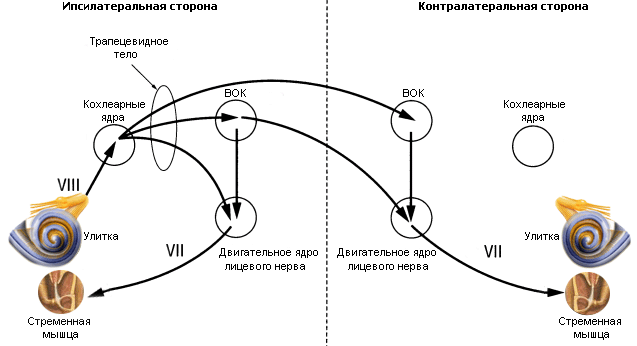
Дуга акустического рефлекса Афферентной (сенсорной) ветвью дуги является слуховой нерв, заканчивающийся в вентральном улитковом ядре, связанном с верхним оливарным комплексом (ВОК) обеих сторон через трапециевидное тело. Эти двусторонние связи обусловливают рефлекторный ответ с обоих ушей, если даже стимулируется только одно ухо. Эфферентный (двигательный) путь дуги простирается от медиального добавочного ядра ВОК до двигательного ядра лицевого нерва (для стременного рефлекса) и двигательного ядра тройничного нерва (для тимпанального рефлекса). Т.о. интенсивный звук (в качестве сенсорного стимула) передается трансформационным аппаратом среднего уха в улитку, где воспринимается слуховым рецептором, кодируется в последовательность нервных импульсов и по слуховому нерву (афферентный центростремительный путь) достигает слуховых центров ствола мозга (ассоциативные центры) – вентральные кохлеарные ядра, верхнеоливарный комплекс (ВОК), трапециевидное тело. Здесь происходит переключение сигнала на двигательные ядра лицевого и тройничного нервов. По ним эфферентные центробежные нервные импульсы достигают соответствующих мышц и вызывают их сокращение.
Патологический процесс может локализоваться на любом участке рефлекторной дуги.
Физиологическое значение АР Вследствие особенностей иннервации, мышца, напрягающая барабанную перепонку, сокращается также при раздражении окончаний тройничного нерва в полости носа, при движении глазных яблок и закрывании глаз, работе некоторых мышц лица и шеи, раздражении воздушной струёй области глазницы, тактильной и электрической стимуляции околоушной области и наружного уха. Причём во всех этих случаях сокращение происходит не изолированно, а сочетании со стременной мышцей. Изолированное сокращение мышцы, напрягающей барабанную перепонку(тимпанальный рефлекс), можно вызвать лишь при электрической стимуляции языка. Порог рефлекса m. stapedii несколько ниже, чем порог рефлекса т. tensoris tympani. Проведенные исследования показали, что латентный период сокращения у m. stapedii оказался более коротким, чем у m. tensoris tympani. При изолированной патологии мышцы, напрягающей барабанную перепонку, рефлекс ещё может регистрироваться, а при поражении стременной мышцы всегда отсутствует. Вообще принято считать, что акустический рефлекс у человека является главным образом, если не исключительно, результатом сокращения стременной мышцы; мышца, напрягающая барабанную перепонку отвечает только на особенно интенсивные звуки. Поэтому иногда АР называют стременным или стапедиальным. АР в норме всегда выявляется с обеих сторон (бинаурально), даже при изолированной стимуляции одного уха. Различают ипсилатеральный АР, когда рефлекс регистрируется в стимулируемом ухе, и контралатеральный рефлекс, выявляемый при стимуляции противоположного уха. Рефлекс может быть вызван звуковым сигналом, подаваемым как через воздух, так и через кость. Сокращение мышц увеличивает жесткость цепи косточек и барабанной перепонки. Стременная мышца тянет головку стремени кнаружи и кзади, а m. tensor tympani — тянет барабанную перепонку внутрь и кпереди, поэтому с первого взгляда может показаться, что они являются антагонистами. Однако действие этих мышц заключается в том, чтобы уменьшить количество энергии, проводимое цепью косточек, и таким образом по отношению к слуху их функции синергичны. Поскольку жесткость обратно пропорциональна частоте (см. механический импеданс), следует ожидать, что АР влияет на процесс передачи звука в среднем ухе в большей степени на низких частотах. Экспериментальные данные подтвердили эти предположения. По данным разных авторов, влияние рефлекса было наибольшим при воздействии звуков с частотой до 2000 Гц (низко- и среднечастотных) и незначительным при звуках более высокой частоты. Пороги восприятия низких звуков при действии АР увеличиваются не менее, чем на 10 дБ (максимально до 45 дБ). Причем это повышение начинает фиксироваться при интенсивности звукового стимула не менее 100 дБ. С повышением интенсивности звука возрастает влияние АР на ослабление входящих акустических сигналов.
Существует много теорий и предположений о значении АР. Поскольку рефлекс регистрируется при относительно высоком уровне стимула, а его величина растет по мере увеличения уровня стимула, следует ожидать, что главное назначение АР состоит в защите улитки от повреждающей стимуляции. Происходит сокращение интратимпанальных мышц и уменьшение колебаний цепи слуховых косточек. Эта «защитная теория» имеет слабые места, так как латентный период и адаптация дают слишком замедленный ответ на внезапно поступающий звук и неэффективны против пролонгированных звуков. Кроме того, звуки, достаточно интенсивные, чтобы вызвать АР, в природе фактически отсутствуют. Тем не менее защита, оказываемая АР, является благотворным моментом, если не главной задачей. Перлманом описана гиперакузия, обусловленная параличом лицевого нерва. Защитная функция мышц барабанной полости показана и экспериментами Като (1913). Кролики, у которых мышцы барабанной полости были перерезаны, под воздействием громкого звука теряли слух быстрее, чем кролики с функционирующими тимпанальными мышцами.
Т.о., рефлекторные сокращения мышц барабанной полости представляют собой «автоматический контроль громкости», правда ограниченной эффективности. Чувствительность уха изменяется в зависимости от интенсивности раздражителя. Сокращение мышц предохраняет внутреннее ухо от чрезмерных звуковых раздражителей. С другой стороны, для звуков малой интенсивности, которые не вызывают сокращения мышц, чувствительность остается высокой. «Аккомодационная теория» рассматривает мышцы как механизм, благодаря которому звукопроводящий аппарат как бы приспосабливается для максимальной передачи звуковой энергии. Согласно «фиксирующей теории», внутрибарабанные мышцы способствуют удержанию косточек в правильном положении и соответствующей ригидности, особенно при действии звуками высокой частоты, когда ускорение звуковых колебаний большое. По данным Simmons тонус мышц среднего уха сглаживает частотный ответ кондуктивной системы. Также он полагает, что модуляции мышечного тонуса повышают «слуховое внимание» путем изменения интенсивности и частотных характеристик окружающих звуков. Эта модуляция аналогична постоянному движению наружных глазных мышц, участвующих в зрении. Поскольку рефлекс главным образом ослабляет низкочастотные звуки и поскольку большинство из собственных физиологических звуков организма — низкие по частоте, АР должен способствовать уменьшению внутреннего шума живого организма. Благодаря снижению маскирующего действия низкочастотных звуков, происходит улучшение восприятия высоких частот, что повышает динамический диапазон слуховой системы.
Рефлекс можно зафиксировать различными методами. Анатомическое положение сухожилия стремени — позади верхнезаднего квадранта pars tensa. Когда перфорация локализуется в этой области, сухожилие мышцы стремени становится непосредственно видимым. Для того, чтобы более ясно наблюдать движение сухожилия рекомендуется небольшое увеличение. Метод непосредственной визуальной регистрации сокращений стременной мышцы через перфорированную барабанную перепонку имеет ряд существенных ограничений. Во-первых, необходимо собственно наличие перфорации, во-вторых, визуально оценить сокращения m. stapedius из-за особенностей анатомического строения бывает непросто, в-третьих, затруднена количественная оценка результатов. Т.о. данный способ весьма субъективен.
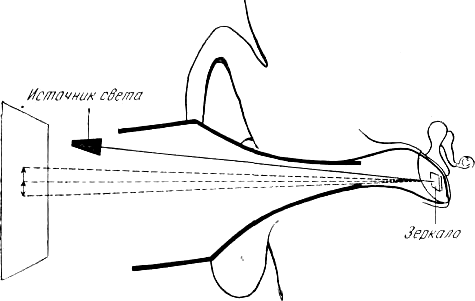
Исследования функции мышц барабанной полости у человека in vivo фактически являются исследованиями вторичного действия мышц на барабанную перепонку. Генрих Кобрак (1947) прикреплял на барабанную перепонку маленькое зеркало и с его помощью регистрировал движения барабанной перепонки, вызванные сокращениями мышц.
Terkildsen также исследовал активность мышц среднего уха непрямым методом, наблюдая изменения воздушного давления в наружном слуховом проходе в ответ на стимуляцию звуком. Сокращение стременной мышцы приводит к смещению барабанной перепонки кнаружи, в то время как сокращение напрягающей барабанную перепонку мышцы смещает ее внутрь. Движения барабанной перепонки в свою очередь приводят к изменениям давления в наружном слуховом проходе. Terkildsen, таким образом, мог судить о природе мышечной активности путем наблюдения за воздушным давлением в слуховом проходе во время звуковой стимуляции. В большинстве его исследований было обнаружено выпячивание барабанной перепонки, что предполагает большую активность стременной мышцы. На сегодняшний день основное значение при регистрации акустического рефлекса (АР) имеют измерения акустического импеданса на поверхности барабанной перепонки с использованием импедансного моста. Механический мост при изучении АР впервые был применен Отто Метцем в Дании, а усовершенствован и внедрен в клиническую практику Джозефом Звислоцким в США. Позже Terkildsen и Nielsen разработали электроакустический импедансный мост. С тех пор почти во всех исследованиях АР стали использовать именно этот метод. Принцип его простой: поскольку сокращение внутрибарабанных мышц делает систему среднего уха жесткой, то импеданс увеличивается (рефлекс в основном действует на компонент жесткости импеданса). Это изменение акустического импеданса и измеряется с помощью моста. Все данные, приведенные ниже, будут касаться АР, зафиксированного посредством измерения акустического импеданса. Параметры АР
Различают ряд количественных параметров АР. Они имеют важное практическое значение, поскольку одновременно являются и диагностическими критериями.
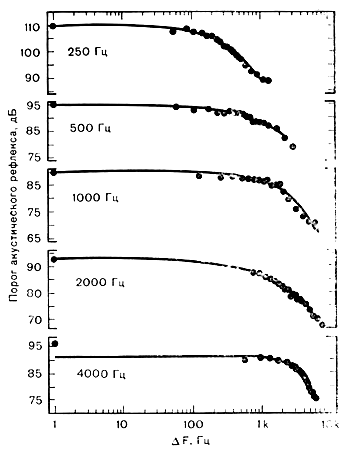
1) Порог АР АР появляется в ответ на звуковую стимуляцию. Минимальная интенсивность звука, способная вызвать сокращение стременной мышцы, которое можно зафиксировать с помощью импедансометрии, является порогом АР. Он рассчитывается в дБ нПС 20 мкПа или УЗД. По данным разных исследователей порог АР в ответ на чистые тоны от 250 до 4000 Гц находится в пределах 80—90 дБ УЗД[9]. Порог рефлекса ниже, когда возбуждающим стимулом служит широкополосный шум, в среднем он составляет 75±5 дБ. Более низкий порог рефлекса для шума, чем для чистого тона, предполагает, что АР имеет отношение к широкому спектру частот стимула. Flottrop и соавт. исследовали эту зависимость, измеряя пороги АР, вызванные широкополосными шумами и сложными тонами. Они обнаружили, что расширение полосы частот не вызывает отклонения величины порога от его значения при стимуляции чистым тоном до тех пор, пока не будет превзойдена определенная полоса частот. В этой точке происходит перелом, после которого расширение полосы частот приводит к более низкому порогу рефлекса. Т.о. существует критическая полоса для АР, выше которой расширение полосы спектра приводит к более низким порогам. Ширина критических полос увеличивается параллельно увеличению центральной частоты стимуляции. Важно заметить, что критическая полоса частот для АР значительно шире, чем психоакустические критические полосы.
Ширина критических полос для порогов АР на частотах 250—4000 Гц. Черные кружки слева соответствуют порогу рефлекса в ответ на воздействие чистых тонов (Popelka et al.)
Наиболее низкие пороги регистрируются при бинауральной стимуляции. Они повышаются на 3 – 6 дБ при ипси- и на 6 – 12 дБ при контралатеральной стимуляции (Brooks D., 1974). Разница между порогами ипси- и контралатеральных АР в зависимости от частоты составляет от 2 до 10 дБ, в среднем 5 дБ (т.е. при ипсилатеральной стимуляции АР вызывается при меньшей интенсивности стимула, чем при контралатеральной). Различают абсолютный и относительный пороги АР. Всё вышесказанное относилось к абсолютному порогу. Относительным порогом АР называют по сути разницу между абсолютным порогом АР и порогом слышимости у данного пациента (на частоте регистрации АР). Он измеряется в дБ над ПЧ пациента. Этот показатель используется при выявлении рекруитмента по данным акустической рефлексометрии (см. ниже).
2) Латентный период АР Акустический рефлекс возникает не одновременно с активирующим сигналом. Время между подачей звукового стимула и началом сдвига импеданса называют латентным периодом АР. Его продолжительность зависит как от интенсивности, так и от частоты стимула. Metz обнаружил, что латентный период уменьшается от 150 мс до 40 мс при повышении уровня сигнала 1000 Гц от 80 дБ до 100 дБ над порогом слуха. Moller сообщил, что латентный период имеет диапазон от 25 до 130 мс для чистых тонов 500 и 1500 Гц. Как правило, латентный период был короче при 1500 Гц, нежели при 500 Гц. Латентный период АР короче при шумовых сигналах (порядка 20 мс), чем при чистых тонах. Эти данные получены при измерениях акустического импеданса. Они характеризуют скорее величину латентного периода механического ответа среднего уха, чем время нервной передачи по рефлекторной дуге. Электромиографический ответ стременной мышцы у человека имеет латентный период всего лишь 12 мс, а электромиографический «порог» АР ниже примерно на 6 дБ. Hung и Dallos сообщили, что у некоторых обследуемых выявлялась «релаксация латентного периода» в начале АР, т.е. перед увеличением импеданса происходило его снижение. Очевидно, этот факт отражает частичную релаксацию стременной мышцы перед ее сокращением.
3) Нарастание (повышение амплитуды) АР Амплитуда АР определяется величиной сдвига импеданса в момент подачи стимула и зависит от интенсивности звука. Увеличение силы подаваемого звука вызывает повышение амплитуды АР.
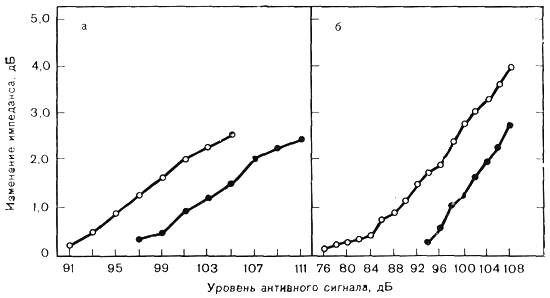
Нарастание рефлекса как функция уровня звукового давления для частоты 1000 Гц (а) и широкополосного шума (б) (Silman et al.). Светлые кружки — у здоровых, темные — у больных.
Соотношение между интенсивностью стимула и амплитудой АР называется функцией нарастания рефлекса. Повышение амплитуды АР происходит линейно для чистых тонов примерно до 120 дБ УЗД. Для широкополосного шума функции достаточно линейны приблизительно до 110 дБ УЗД. На более высоких уровнях УЗД наблюдается насыщение рефлекторного ответа, линейность нарушается. При дальнейшем увеличении интенсивности сигнала можно наблюдать уменьшение амплитуды рефлекса. Flottrop и соавт. установили б о льшие изменения импеданса на частоте 250 Гц, чем на 4000 Гц. Другие исследователи установили, что наибольшие изменения импеданса вызывает тон 2000 Гц; также есть данные, что максимальные ответы вызываются тоном 1000 Гц и широкополосным шумом. Поэтому четкое соотношение между активирующей частотой и амплитудой АР остается до конца нерешенным моментом.
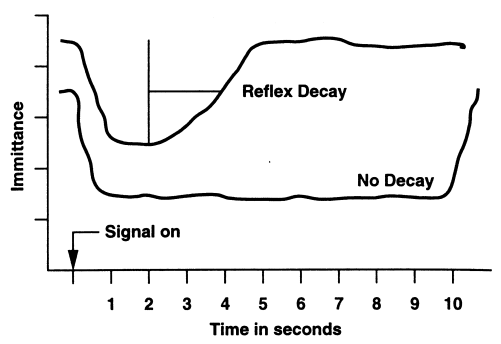
Временные параметры АР Временная суммация относится к зависимости между длительностью стимуляции и интенсивностью, когда время наблюдения составляет менее 1 с. Это легче представить на примере. Предположим, что у обследуемого порог для тона продолжительностью 200 мс оказался равным 18 дБ. Сохранится ли порог на 18 дБ, если тот же тон будет воздействовать в течение лишь 20 мс? Установлено, что если подавать тон длительностью 20 мс, порог изменится до 28 дБ. Это подтверждает общее положение психоакустических исследований, заключающееся в том, что когда сигнал укорачивается в 10 раз (например, с 200 до 20 мс), уровень его должен быть увеличен на 10 дБ, чтобы возместить 10-кратное снижение длительности. По данным Б. С. Мороза (1977), при уменьшении длительности стимула от 1000 до 10 мс, пороги АР повышаются в среднем на 30 дБ. Зависимость между временем действия и интенсивностью стимула условно называется временно-интенсивностной зависимостью. Временная суммация наблюдается как в отношении порога, так и величины АР. Однако оказалось, что изменение интенсивности, необходимое для противодействия заданному снижению длительности стимула, для АР больше. Снижение длительности тона в 2000 Гц со 100 до 10 мс возмещается, по сообщению разных исследователей, увеличением уровня стимула примерно на 15-25 дБ. Величина временной интеграции увеличивается с увеличением частоты, что является общим во всех исследованиях. Рассмотрим стимулы продолжительностью в несколько секунд и более. По мере того как стимуляция продолжается, степень мышечного сокращения и амплитуда АР постепенно снижаются до нулевого уровня,. Это явление называется затуханием (распадом) рефлекса или адаптацией. Сохранение амплитуды ответа на одном уровне в норме можно наблюдать как минимум на протяжении 10-15 с. Затем амплитуда АР начинает постепенно понижаться, причём тем быстрее, чем выше частота стимула. Поэтому оценивать явление неустойчивости амплитуды, обозначаемое термином «распад рефлекса», нужно исходя из частоты стимулирующего сигнала. При широкополосном шуме время действия АР максимально – 120-150 с. Адаптация рефлекса увеличивается по мере роста частоты тонального стимула: для тона 500 Гц время действия АР в среднем равно 120 с, для тона 2000 Гц – 44 с, а для тона 4000 Гц всего 16 с.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 66; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.55.42 (0.033 с.) |