Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гормоны. Химическая природа. Классификация. Связь структуры гормонов с механизмом их действия.Стр 1 из 17Следующая ⇒
Гормоны. Химическая природа. Классификация. Связь структуры гормонов с механизмом их действия. К гормонам относят разнообразные по химической природе соединения, вырабатываемые в эндокринных железах, секретируемые непосредственно в кровь, оказывающие дистанционный биологический эффект. Они являются гуморальными посредниками, которые обеспечивают поступление сигнала в клетки-мишени и вызывают специфические изменения в сенситивных к ним тканях и органах. Гормоны оказывают свой эффект в очень малых концентрациях (10-3–10-12 моль/л). У каждого из них существует свой ритм секреции в течение суток, месяца или времени года, специфический для каждого гормона период жизни, как правило, очень короткий (секунды, минуты, редко часы). По химической природе гормональные молекулы относят к трем группам соединений: - белки и пептиды; - производные аминокислот; - стероиды и производные жирных кислот. --- Гормоны задней доли гипофиза: окситоцин, вазопрессин. Их химическая природа и биологическое действие Окситоцин Строение Представляет собой пептид, состоящий из 9 аминокислот и периодом полураспада 5 минут. Синтез Осуществляется в гипоталамусе. В точку секреции – заднюю долю гипофиза – гормон попадает по аксонам с белком-переносчиком нейрофизином. Регуляция синтеза и секреции Стимулируют секрецию раздражение грудных сосков (кормление грудью), беременность, увеличение частоты секс-контактов, стресс, сон, серотонин. Механизм действия Точно не известен. Связан с регуляцией количества простагландинов в клетке и изменением потоков ионов Cа2+ и Na+. Мишени и эффекты Матка Изменяя ионные потоки в миометрии матки, вызывает ее сокращение. С повышением срока беременности чувствительность матки к гормону возрастает. Молочная железа В миоэпителиальных клетках альвеол стимулирует спазм протоков и выделение молока. Жировая ткань Увеличивает потребление глюкозы и, следовательно, синтез триацилглицеролов. Патология не отмечено Вазопрессин Строение Представляет собой пептид, включающий 9 аминокислот, с периодом полураспада 2-4 минуты. Синтез Осуществляется в супраоптическом и паравентрикулярном ядрах гипоталамуса. Отсюда в точку секреции (заднюю долю гипофиза) вазопрессин отправляется в виде прогормона, состоящего из двух частей – собственно АДГ и нейрофизина. В ходе транспортировки происходит процессинг – гидролиз проАДГ на зрелый гормон и белок нейрофизин.
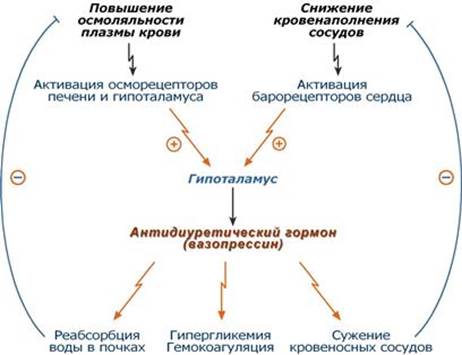
Регуляция синтеза и секреции Уменьшают: этанол, глюкокортикоиды. Активируют: - возбуждение осморецепторов в гипоталамусе и в портальной вене печени из-за повышения осмолярности плазмы при обезвоживании, почечной или печеночной недостаточности, накоплении осмотически активных веществ (глюкоза), - активация барорецепторов сердца и каротидного синуса при снижении объема крови в сосудистом русле (кровопотери, обезвоживание), - эмоциональный и физический стресс, - никотин, ангиотензин II, интерлейкин 6, морфин, ацетилхолин.
Механизм действия Зависит от рецепторов: 1. Кальций-фосфолипидный механизм, сопряжен: - с V1-рецепторами гладких мышц артериол, печени, тромбоцитов, - с V3-рецепторами аденогипофиза и структур головного мозга. 2. Аденилатциклазный механизм – с V2-рецепторами почечных канальцев. Мишени и эффекты Почки Увеличивает реабсорбцию воды в эпителиоцитах дистальных канальцев и собирательных трубочек, благодаря "выставлению" на мембрану транспортных белков для воды – аквапоринов: - через аденилатциклазный механизм вызывает фосфорилирование молекул аквапоринов (только тип 2, AQP2), их взаимодействие с белками микротубул и путем экзоцитоза встраивание аквапоринов в апикальную мембрану, - по тому же механизму стимулирует синтез аквапоринов de novo. Сосудистая система Поддерживает стабильное давление крови, стимулируя тонус сосудов: - повышает тонус гладких мышц сосудов кожи, скелетных мышц и миокарда (в меньшей степени), - повышает чувствительность механорецепторов в каротидных синусах к изменениям артериального давления, --- 4. Гормоны щитовидной железы. Их строение и образование. Механизм действия. Гипо- и гипертиреоз. Тиреотропный гормон Строение Представляет собой гликопротеин с молекулярной массой 30 кДа, состоит из двух субъединиц α- и β, α-субъединица схожа с таковой гонадотропных гормонов, β-субъединица специфична для ТТГ.
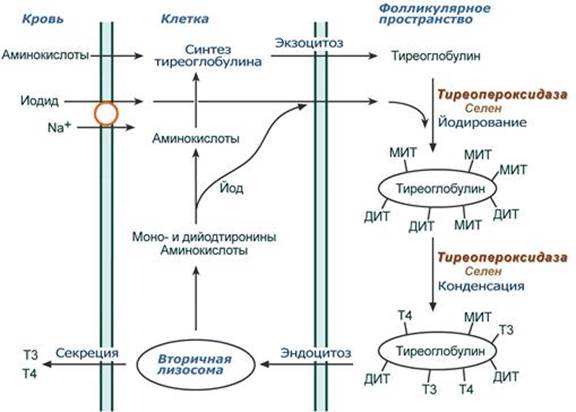
Синтез Осуществляется в базофильных тиреотрофах гипофиза. Механизм действия Аденилатциклазный, связанный с ингибированием GI-белка и накоплением цАМФ, и кальций-фосфолипидный механизм с образованием инозитол-трифосфата, диацилглицерола и комплекса кальций-кальмодулин. Мишени и эффекты Тиреотропный гормон в щитовидной железе: 1. Обеспечивает жизнедеятельность щитовидной железы - стимулирует углеводный обмен (гликолиз, ПФП), синтез гетерополисахаридов - повышает синтез белков, фосфолипидов и нуклеиновых кислот, - стимулирует васкуляризацию щитовидной железы - стимулирует рост и пролиферацию тиреоидных клеток 2. Стимулирует гормональную активность щитовидной железы - повышает захват йода и его включение в тиреоглобулин - активирует все стадии образования трийодтиронина и тироксина, в том числе увеличивает экспрессию гена тиреопероксидазы. Патология При уменьшении выработки изменяется масса тела, повышается утомляемость, возникают симптомы гипотиреоза. Йодтиронины Строение К гормонам самой щитовидной железы относятся тироксин и трийодтиронин, которые представляют собой йодированные производные аминокислоты тирозина. Синтез Осуществляется в фолликулярных клетках щитовидной железы. Йодиды поступают из крови в клетку симпортом с ионами Na+ и из клетки в фолликулярное пространство диффузией. На апикальной мембране клеток селен-зависимая гемсодержащая тиреопероксидаза: - йодирует остатки тирозина в тиреоглобулине с образованием моно- и дийодпроизводных (МИТ, ДИТ) тирозина, - конденсирует часть МИТ и ДИТ до йодтиронинов, при этом доля трийодтиронина (Т3) и тетрайодтиронина (тироксин, Т4) составляет около 30% от всех йодпроизводных.
Йодированный тиреоглобулин хранится во внеклеточных коллоидах, при тиреотропной стимуляции пиноцитируется фолликулярными клетками, сливается с лизосомами и гидролизуется. Далее три- и тетрайодтиронин секретируются в кровь. В крови гормоны транспортируются специфическим глобулином, а также альбумином. Механизм действия Цитозольный. Мишени и эффекты Рецепторы к йодтиронинам имеют все ткани организма. В клетках-мишенях, особенно в печени, тироксин дейодируется и активной формой является трийодтиронин (3,5,3'-производное). --- Кальцитриол
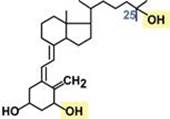
Строение Представляет собой производное витамина D и относится к стероидам. Синтез Образующийся в коже под действием ультрафиолета и поступающие с пищей холекальциферол (витамин D3) и эргокальциферол (витамин D2) гидроксилируются в гепатоцитах по С25 и в эпителии проксимальных канальцев почек по С1. В результате формируется 1,25-диоксихолекальциферол (кальцитриол). Механизм действия Цитозольный. Мишени и эффекты Рецепторы к кальцитриолу обнаружены практически во всех тканях. Эффекты гормона могут быть геномные и негеномные, эндокринные и паракринные. 1. Геномный эффект кальцитриола заключается в регуляции концентрации кальция и фосфора в крови: - в кишечнике индуцирует синтез белков, отвечающих за всасывание кальция и фосфатов - в почечных канальцах повышает синтез белков-каналов для реабсорбции ионов кальция и фосфатов
- в костной ткани усиливает активность остеобластов и остеокластов. 2. Посредством негеномных паракринных механизмов гормон регулирует количество ионов Ca2+ в клетке: - способствует минерализации кости остеобластами, - влияет на активность иммунных клеток, модулируя их иммунные реакции, - участвует в проведении нервного возбуждения, - поддерживает тонус сердечной и скелетной мускулатуры, - влияет на пролиферацию клеток. 3. Также действие кальцитриола сопровождается подавлением секреции паратиреоидного гормона. Паратиреоидный гормон Строение Представляет собой пептид из 84 аминокислот с молекулярной массой 9,5 кДа. Синтез Идет в паращитовидных железах. Реакции синтеза гормона высоко активны.
Механизм действия Аденилатциклазный. Мишени и эффекты Эффект паратиреоидного гормона заключается в увеличении концентрации кальция и снижении концентрации фосфатов в крови. Это достигается тремя способами: Костная ткань - при высоком уровне гормона активируются остеокласты и происходит деструкция костной ткани - при низких концентрациях активируется перестройка кости и остеогенез. Почки - увеличивается реабсорбция кальция и магния - уменьшается реабсорбция фосфатов, аминокислот, карбонатов, натрия, хлоридов, сульфатов. - также гормон стимулирует образование кальцитриола (гидроксилирование по С1). Кишечник - при участии кальцитриола усиливается всасывание кальция и фосфатов. Гипофункция Возникает при случайном удалении железы при операциях на щитовидной железе или при аутоиммунной деструкции ткани желез. Возникающая гипокальциемия и гиперфосфатемия проявляется в виде высокой нервно-мышечной возбудимости, судорог, тетании. При резком снижении кальция возникает дыхательный паралич, ларингоспазм. Гиперфункция Первичный гиперпаратиреоз возникает при аденоме желез. Нарастающая гиперкальциемия вызывает повреждение почек, мочекаменную болезнь. Вторичный гиперпаратиреоз является результатом почечной недостаточности, при которой происходит нарушение образования кальцитриола, снижение концентрации кальция в крови и компенсаторное возрастание синтеза паратиреоидного гормона. Кальцитонин Строение Представляет собой пептид, включающий 32 аминокислоты с молекулярной массой 3,6 кДа.
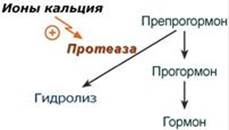
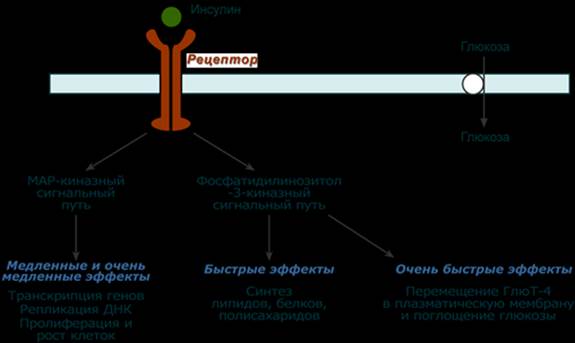
Синтез Осуществляется в парафолликулярных клетках щитовидной железы. Механизм действия Аденилатциклазный Мишени и эффекты Эффект кальцитонина заключается в уменьшении концентрации кальция и фосфатов в крови: - в костной ткани подавляет активность остеокластов, что улучшает вход кальция и фосфатов в кость - в почках подавляет реабсорбцию ионов Ca2+, фосфатов, Na+, K+, Mg2+. --- Строение Инсулин представляет собой белок, состоящий из двух пептидных цепей А (21 аминокислота) и В (30 аминокислот), связанных между собой дисульфидными мостиками. Всего в зрелом инсулине человека присутствует 51 аминокислота и его молекулярная масса равна 5,7 кДа. Синтез Инсулин синтезируется в β-клетках поджелудочной железы в виде препроинсулина, на N-конце которого находится концевая сигнальная последовательность из 23 аминокислот, служащая проводником всей молекулы в полость эндоплазматической сети. Здесь концевая последовательность сразу отщепляется и проинсулин транспортируется в аппарат Гольджи. Мишени Рецепторы инсулина находятся практически на всех клетках организма, кроме нервных, но в разном количестве. Нервные клетки не имеют рецепторов к инсулину, т.к. последний просто не проникает через гематоэнцефалический барьер. Механизм действия После связывания инсулина с рецептором активируется ферментативный домен рецептора. Так как он обладает тирозинкиназной активностью, то фосфорилирует внутриклеточные белки - субстраты инсулинового рецептора. Дальнейшее развитие событий обусловлено двумя направлениями: MAP-киназный путь и ФИ-3-киназный механизмы действия. При активации фосфатидилинозитол-3-киназного механизма результатом являются быстрые эффекты – активация ГлюТ-4 и поступление глюкозы в клетку, изменение активности "метаболических" ферментов – ТАГ-липазы, гликогенсинтазы, гликогенфосфорилазы, киназы гликогенфосфорилазы, ацетил-SКоА-карбоксилазы и других. При реализации MAP-киназного механизма (англ. MAP - mitogen-activated protein) регулируются медленные эффекты – пролиферация и дифференцировка клеток, процессы апоптоза и антиапоптоза.
Быстрые эффекты (минуты) Быстрые эффекты заключаются в изменении скоростей фосфорилирования и дефосфорилирования метаболических ферментов и регуляторных белков. Печень - торможение эффектов адреналина и глюкагона (фосфодиэстераза) - ускорение гликогеногенеза (гликогенсинтаза) - активация гликолиза (фосфофруктокиназа, пируваткиназа) - превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа) - усиление синтеза жирных кислот (ацетил-SКоА-карбоксилаза) - формирование ЛПОНП - повышение синтеза холестерина (ГМГ-SКоА-редуктаза) Мышцы - торможение эффектов адреналина (фосфодиэстераза) - стимулирует транспорт глюкозы в клетки (активация ГлюТ-4) - стимуляция гликогеногенеза (гликогенсинтаза) - активация гликолиза (фосфофруктокиназа, пируваткиназа) - превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа)
- усиливает транспорт нейтральных аминокислот в мышцы - стимулирует трансляцию (рибосомальный синтез белков) Жировая ткань - стимулирует транспорт глюкозы в клетки (активация Глют-4) - активирует запасание жирных кислот в тканях (липопротеинлипаза) - активация гликолиза (фосфофруктокиназа, пируваткиназа) - усиление синтеза жирных кислот (активация ацетил-SКоА-карбоксилазы) - создание возможности для запасания ТАГ (инактивация гормон-чувствительной-липазы) Глюкагон Строение Представляет собой полипептид, включающий 29 аминокислот с молекулярной массой 3,5 кДа и периодом полураспада 3-6 мин. Синтез Осуществляется в клетках поджелудочной железы и в клетках тонкого кишечника. Механизм действия Аденилатциклазный активирующий. Мишени и эффекты Конечным эффектом является повышение концентрации глюкозы и жирных кислот в крови. Жировая ткань Повышает активность внутриклеточной гормон-чувствительной ТАГ-липазы и, соответственно, стимулирует липолиз. Печень - активация глюконеогенеза и гликогенолиза, - за счет повышенного поступления жирных кислот из жировой ткани усиливает кетогенез. Патология Гиперфункция Глюкагонома – редко встречающееся новообразование из группы нейроэндокринных опухолей. У больных отмечается гипергликемия и поражение кожи и слизистых оболочек. --- Строение Глюкокортикоиды являются производными холестерола и имеют стероидную природу. Основным гормоном у человека является кортизол.
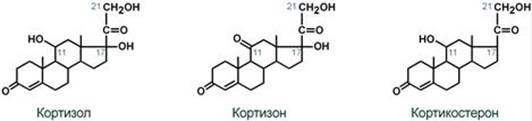
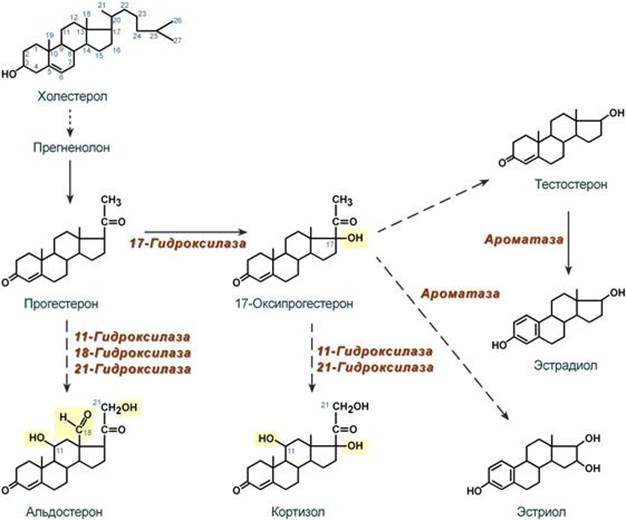
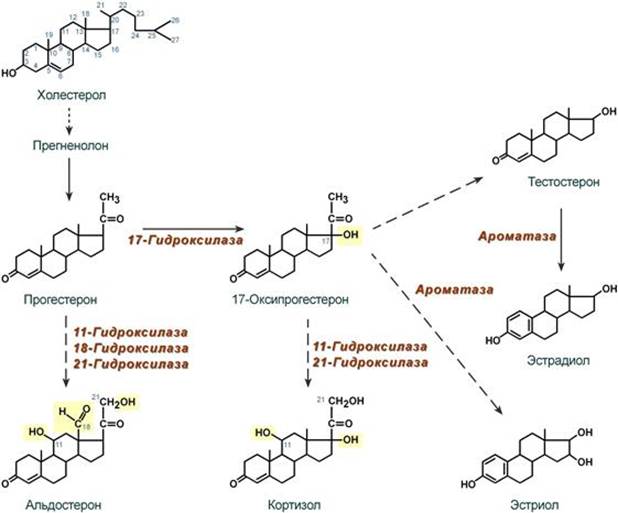
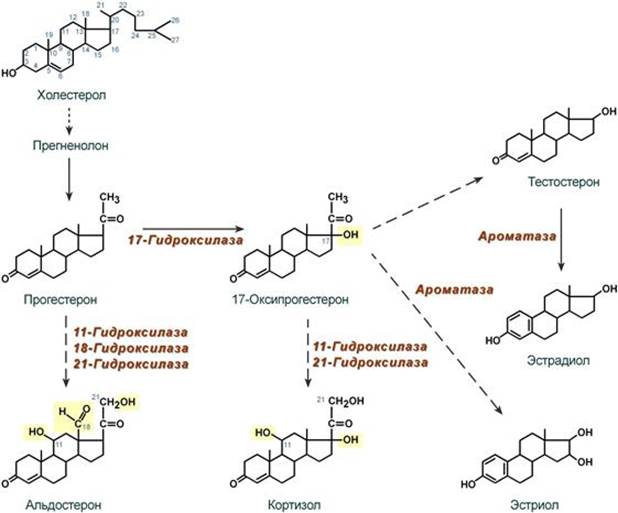
Синтез Осуществляется в сетчатой и пучковой зонах коры надпочечников. Образованный из холестерола прогестерон подвергается окислению 17-гидроксилазой по 17 атому углерода. После этого в действие последовательно вступают еще два значимых фермента: 21-гидроксилаза и 11-гидроксилаза. В конечном итоге образуется кортизол.
Механизм действия Цитозольный. Мишени и эффекты Мишенью является лимфоидная, эпителиальная (слизистые оболочки и кожа), жировая, костная и мышечная ткани, печень. Белковый обмен - значительное повышение катаболизма белков в лимфоидной, эпителиальной, мышечной, соединительной и костной тканях, - в печени в целом стимулирует анаболизм белков (например, ферменты трансаминирования и глюконеогенеза), - стимуляция реакций трансаминирования через синтез аминотрансфераз, обеспечивающих удаление аминогрупп от аминокислот и получение углеродного скелета кетокислот, Углеводный обмен В целом вызывают повышение концентрации глюкозы крови: - усиление мощности глюконеогенеза из кетокислот за счет увеличения синтеза фосфоенолпируват-карбоксикиназы, - увеличение синтеза гликогена в печени за счет активации фосфатаз и дефосфорилирования гликогенсинтазы. - снижение проницаемости мембран для глюкозы в инсулинзависимых тканях. Липидный обмен - стимуляция липолиза в жировой ткани благодаря увеличению синтеза ТАГ-липазы, что усиливает эффект АКТГ, СТГ, глюкагона, катехоламинов, т.е. кортизол оказывает пермиссивное действие (англ. permission - позволение). Водно-электролитный обмен - слабый минералокортикоидный эффект на канальцы почек вызывает реабсорбцию натрия и потерю калия, - потеря воды в результате подавления секреции вазопрессина и излишняя задержка натрия из-за увеличения активности ренин-ангиотензин-альдостероновой системы. --- Синтез Осуществляется в клубочковой зоне коры надпочечников. Образованный из холестерола прогестерон на пути к альдостерону подвергается последовательному окислению 21-гидроксилазой, 11-гидроксилазой и 18-гидроксилазой. В конечном итоге образуется альдостерон.
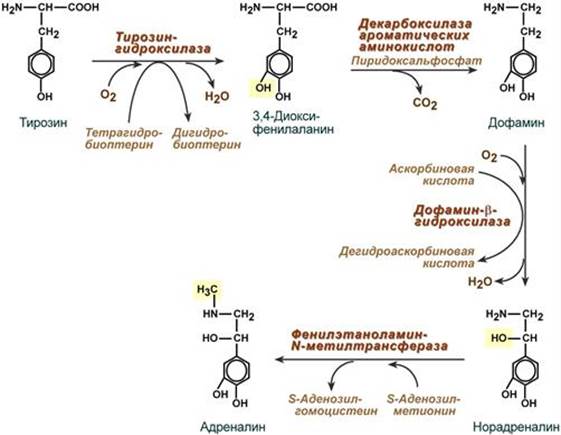
Механизм действия Цитозольный. Мишени и эффекты Воздействует на слюнные железы, на дистальные канальцы и собирательные трубочки почек. В почках усиливает реабсорбцию ионов натрия и потерю ионов калия посредством следующих эффектов: - увеличивает количество Na+,K+-АТФазы на базальной мембране эпителиальных клеток, - стимулирует синтез митохондриальных белков и увеличение количества нарабатываемой в клетке энергии для работы Na+,K+-АТФазы, - стимулирует образование Na-каналов на апикальной мембране клеток почечного эпителия. --- Строение Гормоны надпочечников адреналин и норадреналин под общим названием катехоламины представляют собой производные аминокислоты тирозина. Роль адреналина является гормональной, норадреналин преимущественно является нейромедиатором. Синтез Осуществляется в клетках мозгового слоя надпочечников (80% всего адреналина), синтез норадреналина (80%) происходит также в нервных синапсах.
Механизм действия Механизм действия гормонов разный в зависимости от рецептора. Степень активности рецептора может изменяться в зависимости от концентрации соответствующего лиганда. Например, в жировой ткани при низких концентрациях адреналина более активны α2-адренорецепторы, при повышенных концентрациях (стресс) – стимулируются β1-, β2-, β3-адренорецепторы. Кальций-фосфолипидный механизм - при возбуждении α1-адренорецепторов. Аденилатциклазный механизм - при задействовании α2-адренорецепторов аденилатциклаза ингибируется, - при задействовании β1- и β2-адренорецепторов аденилатциклаза активируется. Мишени и эффекты α1-Адренорецепторы При возбуждении α1-адренорецепторов происходит: 1. Активация гликогенолиза и глюконеогенеза в печени. 2. Сокращение гладких мышц: - кровеносных сосудов в разных областях тела, - мочеточников и сфинтера мочевого пузыря, - предстательной железы и беременной матки, - радиальной мышцы радужной оболочки, - поднимающих волос, - капсулы селезенки. 3. Расслабление гладких мышц ЖКТ и сокращение его сфинктеров, α2-Адренорецепторы При возбуждении α2-адренорецепторов происходит: - снижение липолиза в результате уменьшения стимуляции ТАГ-липазы, - подавление секреции инсулина и секреции ренина, - спазм кровеносных сосудов в разных областях тела, - расслабление гладких мышц кишечника, - стимуляция агрегации тромбоцитов. β1-Адренорецепторы Возбуждение β1-адренорецепторов (есть во всех тканях) проявляется в основном: - активация липолиза, - расслабление гладких мышц трахеи и бронхов, - расслабление гладких мышц ЖКТ, - увеличение силы и частоты сокращений миокарда (ино- и хронотропный эффект). β2-Адренорецепторы Возбуждение β2-адренорецепторов (есть во всех тканях) проявляется главным образом: 1. Стимуляция: - гликогенолиза и глюконеогенеза в печени, - гликогенолиза в скелетных мышцах, - липолиза в жировой ткани. 2. Усиление секреции: - инсулина, - тиреоидных гормонов. 3. Расслабление гладких мышц: - трахеи и бронхов, - желудочно-кишечного тракта, - беременной и небеременной матки, - кровеносных сосудов в разных областях тела, - мочеполовой системы, - капсулы селезенки, 4. Усиление сократительной активности скелетных мышц (тремор), 5. Подавление выхода гистамина из тучных клеток. --- Тестостерон Строение По строению являются производными холестерола, т.е. стероидами.
Синтез Мужские гормоны: на 95% тестостерон синтезируется в клетках Лейдига семенников и на 5% в периферических тканях при метаболизме андрогенов, образованных в сетчатой и пучковой зонах надпочечников.
Механизм действия Цитозольный. Мишени и эффекты Тестостерон вызывает синтез всего, что позволяет увеличить массу тканей. Биохимические эффекты: - задержка азота, - активация синтеза белков, РНК, ДНК, липидов, полисахаридов большинства тканей. Другие эффекты: - совместно с СТГ ускоряет рост кости в отрочестве с последующей остановкой роста и закрытием эпифизов, - усиливает продукцию эритроцитов, - обусловливает рост мышц, - стимулирует сальные железы, что повышает жирность кожи и вызывает возникновение угрей (acne vulgaris). Может наблюдаться у подростков обоего пола и у женщин в менопаузе, - увеличивает общую пигментацию, снижение тембра голоса, - определяет распределение жира, - увеличивает сперматогенез и развитие предстательной железы, - отвечают за сексуальное влечение (либидо) как у мужчин, так и у женщин. --- Строение Являются производными холестерола – стероиды.
Женские гормоны: эстрогены синтезируются в фолликулах яичников, прогестерон – в желтом теле. Частично гормоны могут образовываться в адипоцитах в результате ароматизации андрогенов.
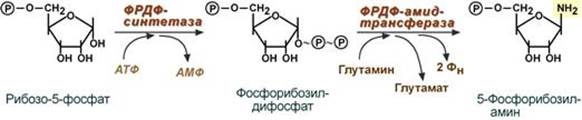
Мишени и эффекты Эстрогены 1. При половом созревании эстрогены активируют синтез белка и нуклеиновых кислот в органах половой сферы и обеспечивают формирование половых признаков: ускоренный рост и закрытие эпифизов длинных костей, определяют распределение жира на теле, пигментацию кожи, стимулируют развитие влагалища, маточных труб, матки, развитие стромы и протоков грудных желез, рост подмышечных и лобковых волос. 2. В организме взрослой женщины: Биохимические эффекты: - активирует в печени синтез транспортных белков для тироксина, железа, меди и т.п., - стимулирует синтез факторов свертывания крови – II, VII, IX, X, плазминогена, фибриногена, подавляет синтез антитромбина III и адгезию тромбоцитов, - увеличивает синтез ЛПВП, подавляет ЛПНП, повышает концентрацию ТАГ в крови и снижает содержание холестерола, - снижает резорбцию кальция из костной ткани. Другие эффекты: - стимулирует рост железистого эпителия эндометрия, - определяет структуру кожи и подкожной клетчатки, - подавляет перистальтику кишечника, что повышает абсорбцию веществ. Прогестерон Прогестерон является основным гормоном второй половины цикла и его задача – обеспечить наступление и сохранение беременности. Биохимические эффекты: - повышает активность липопротеинлипазы на эндотелии капилляров, - увеличивает концентрацию инсулина в крови, - подавляет реабсорбцию натрия в почках, - является ингибитором ферментов дыхательной цепи, что снижает катаболизм, - ускоряет выведение азота из организма женщины. Другие эффекты: - расслабляет мышцы беременной матки, - усиливает реакцию дыхательного центра на СО2, что снижает в крови парциальное давление СО2 при беременности и в лютеиновую фазу цикла, - обусловливает рост молочной железы при беременности, - сразу после овуляции является хематтрактантом для сперматозоидов, движущихся по маточным трубам. --- Биосинтез пуриновых нуклеотидов. Исходные субстраты синтеза. Регуляция синтеза. Роль витаминов в механизмах синтеза. Синтез пуриновых оснований происходит во всех клетках организма, главным образом в печени. Исключение составляют эритроциты, полиморфноядерные лейкоциты, лимфоциты. Условно все реакции синтеза можно разделить на 4 этапа: 1. Синтез 5'-фосфорибозиламина Первая реакция синтеза пуринов заключается в активации углерода в положении С1 рибозо-5-фосфата, это достигается синтезом 5-фосфорибозил-1-дифосфата (ФРДФ). Фосфорибозил-дифосфат является тем якорем, на основе которого синтезируется сложный пуриновый цикл. Вторая реакция – это перенос NH2-группы глутамина на активированный атом С1 с образованием 5'-фосфорибозиламина. Указанная NH2-группа фосфорибозиламина уже принадлежит будущему пуриновому кольцу и ее азот будет атомом номер 9.
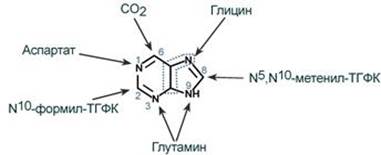
2. Синтез инозинмонофосфата 5-фосфорибозиламин вовлекается в девять реакций, и в результате образуется первый пуриновый нуклеотид – инозинмонофосфорная кислота (ИМФ). В этих реакциях источниками атомов пуринового кольца являются глицин, аспартат, еще одна молекула глутамина, углекислый газ и производные тетрагидрофолиевой кислоты (ТГФК). В целом на синтез пуринового кольца затрачивается энергия 6 молекул АТФ.
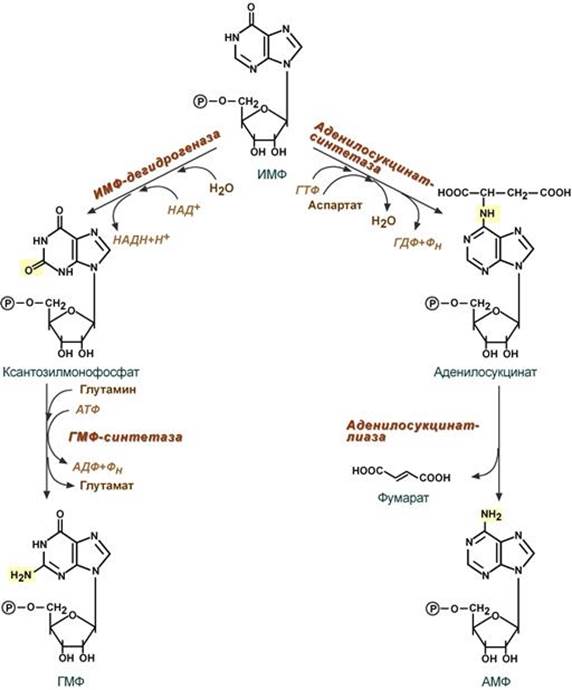
3. Синтез аденозинмонофосфата и гуанозинмонофосфата Гуанозинмонофосфат (ГМФ) образуется в двух реакциях – сначала ИМФ окисляется ИМФ-дегидрогеназой до ксантозилмонофосфата, источником кислорода является вода, акцептором водорода – НАД. После этого работает ГМФ-синтетаза, она использует универсальный клеточный донор NH2-групп – глутамин, источником энергии для реакции служит АТФ. Аденозинмонофосфат (АМФ) также образуется в двух реакциях, но в качестве донора NH2-группы выступает аспарагиновая кислота. В первой, аденилосукцинат-синтетазной, реакции на присоединение аспартата используется энергия распада ГТФ, во второй реакции аденилосукцинат-лиаза производит удаление части аспарагиновой кислоты в виде фумарата.
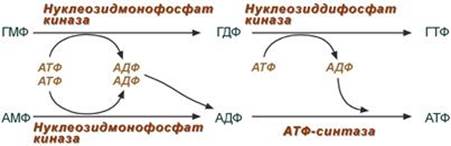
4. Образование нуклеозидтрифосфатов АТФ и ГТФ. Синтез ГТФ осуществляется в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ. Синтез АТФ происходит несколько иначе. АДФ из АМФ образуется также за счет макроэргических связей АТФ. Для синтеза же АТФ из АДФ в митохондриях есть фермент АТФ-синтаза, образующий АТФ в реакциях окислительного фосфорилирования.
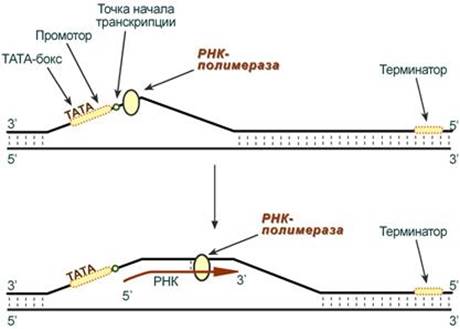
--- Стадии транскрипции Выделяют три стадии транскрипции: инициация, элонгация и терминация. Инициация Промотор содержит стартовый сигнал транскрипции – ТАТА-бокс. Так называется определенная последовательность нуклеотидов ДНК, связывающая первый фактор инициации ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции (матричная нить ДНК). Так как промотор ассиметричен ("ТАТА"), то он связывает РНК-полимеразу только в одной ориентации, что определяет направление транскрипции от 5'-конца к 3'-концу (5'→3'). Для связывания РНК-полимеразы с промотором необходим еще один фактор инициации – σ-фактор (греч. σ – "сигма"), но сразу после синтеза затравочного фрагмента РНК (длиной 8-10 рибонуклеотидов) σ-фактор отрывается от фермента. Другие факторы инициации раскручивают спираль ДНК перед РНК-полимеразой.
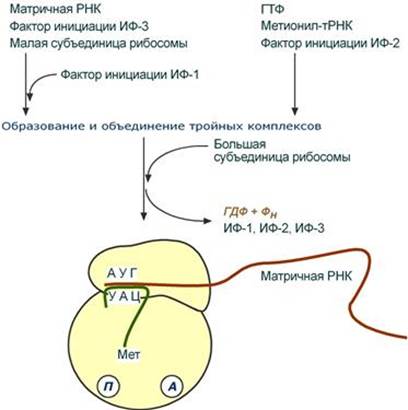
Элонгация Белковые факторы элонгации обеспечивают продвижение РНК-полимеразы вдоль ДНК и расплетают молекулу ДНК на протяжении примерно 17 нуклеотидных пар. РНК-полимераза продвигается со скоростью 40-50 нуклеотидов в секунду в направлении 5'→3'. Фермент использует АТФ, ГТФ, ЦТФ, УТФ одновременно в качестве субстрата и в качестве источника энергии. Терминация РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью белкового фактора терминации, так называемого ρ-фактора (греч. ρ – "ро"), от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая является первичным транскриптом, предшественником мРНК или тРНК или рРНК. --- Инициация Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина. В начале этой стадии формируются два тройных комплекса: - первый комплекс – мРНК + малая субъединица + ИФ-3, - второй комплекс – метионил-тРНК + ИФ-2 + ГТФ. После формирования тройные комплексы объединяются с большой субъединицей рибосомы. В этом процессе активно участвуют белковые факторы инициации, источником энергии служит ГТФ. После сборки комплекса инициирующая метионил-тРНК связывается с первым кодоном АУГ матричной РНК и располагается в П-центре (пептидильный центр) большой субъединицы. А-центр (аминоацильный центр) остается свободным, он будет задействован на стадии элонгации для связывания аминоацил-тРНК.
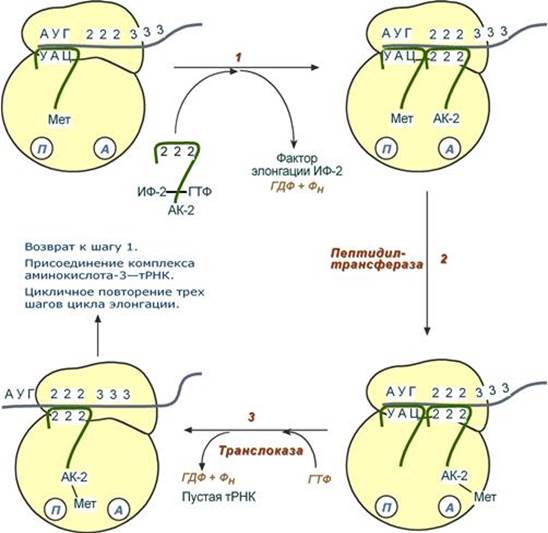
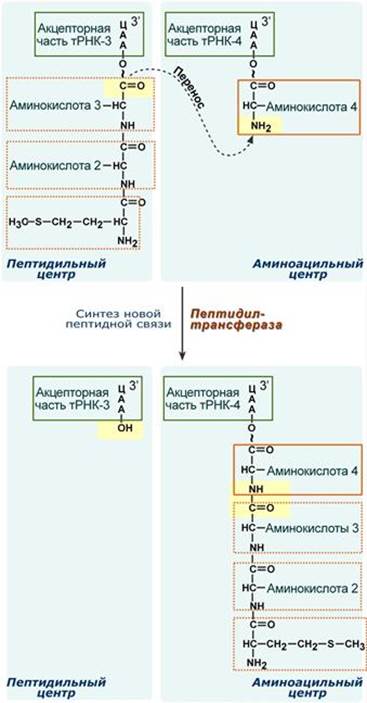
После присоединения большой субъединицы начинается стадия элонгации. Элонгация Для этой стадии необходимы все 20 аминокислот, тРНК для всех аминокислот, белковые факторы элонгации, ГТФ. Удлинение цепи происходит со скоростью примерно 20 аминокислот в секунду. Элонгация представляет собой циклический процесс. Первый цикл (и следующие циклы) элонгации включает три шага: - Присоединение аминоацил-тРНК (еще второй) к кодону мРНК (еще второму), аминокислота при этом встраивается в А-центр рибосомы. Источником энергии служит ГТФ. - Фермент пептидилтрансфераза осуществляет перенос метионина с метионил-тРНК (в П-центре) на вторую аминоацил-тРНК (в А-центре) с образованием пептидной связи между метионином и второй аминокислотой. При этом уже активированная СООН-группа метионина связывается со свободной NH2-группой второй аминокислоты. Здесь источником энергии служит макроэргическая связь между аминокислотой и тРНК. - Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы, второй кодон (на рисунке) становится напротив П-центра, напротив А-центра оказывается третий кодон (на рисунке). Для этих процессов необходима затрата энергии ГТФ. Так как вместе с мРНК перемещаются закрепленные на ней тРНК, то инициирующая первая тРНК выходит из рибосомы, вторая тРНК с дипептидом помещается в П-центр.
Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А-центр, после чего он смещается в П-центр в транслоказной реакции.. В пустой А-центр входит четвертая аминоацил-тРНК и начинается третий цикл элонгации:
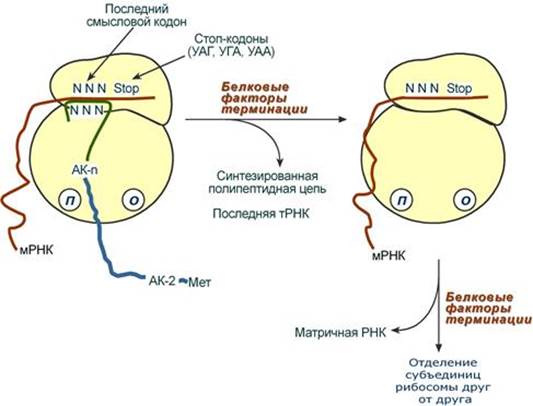
Цикл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь. Терминация Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют: - Гидролитическое отщепление полипептида от конечной тРНК. - Отделение от П-центра последней, уже пустой, тРНК. - Диссоциацию рибосомы. Источником энергии для завершения трансляции является ГТФ.
--- Ингибирование 1. Гетероциклические соединения доксорубицин, дауномицин и актиномицин D обладают способностью интеркалировать (встраиваться между нитей молекулы ДНК) между двумя соседними парами оснований Г-Ц. В результате возникает препятствие для движения РНК-полимеразы ("заедание молнии") и остановка транскрипции. 2. Р ифампицин связывается с β-субъединицей РНК-полимеразы прокариот и ингибирует ее. Благодаря такой избирательности действия рифампицин действует только на бактерии и является препаратом для лечения туберкулеза. 3. α-Аманитин, октапептид бледной поганки (Amanita phalloides) блокирует РНК-полимеразу II эукариот и предотвращает продукцию мРНК. Активация Активация транскрипции используется в клинике намного реже и заключается в применении аналогов стероидных гормонов для достижения анаболического эффекта в органе-мишени. --- Механизмы регуляции биохимических реакций. Обратимая и необратимая регуляция. Представление о механизме изостерической регуляции. Использование принципов изостерической регуляции в медицинской практике Механизмы регуляции биохимических реакций: 1. Регуляция количества ферментов: индукция/репрессия синтеза (стероидные и тиреоидные гормоны индуцируют синтез ферментов), распад ферментов (скорость разрушения ферментов) 2. Регуляция активности ферментов: А) ингибитор Б) аллостерия и кооперативность В) регуляция активности путем ковалентной модификации структуры ферментов Г) регуляция активности при помощи белков-регуляторов 3. Изменение количества субстрата 4. Наличие изоферментов<
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 159; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.174.76 (0.262 с.) |