Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности энергетического обмена в мышцах; креатинфосфат. Биохимические изменения при мышечных дистрофиях и деинервации мышц. Креатинурия.
Прежде всего ресинтез АТФ обеспечивается трансфосфорилированием АДФ с креатинфосфатом. Данная реакция катализируется ферментом креатинкиназой:
Креатинкиназный путь ресинтеза АТФ является чрезвычайно быстрым и максимально эффективным (за счет каждой молекулы креатинфосфата образуется молекула АТФ). Именно поэтому долгое время не удавалось установить уменьшение концентрации АТФ и соответственно повышение концентрации АДФ даже при достаточно продолжительном тетанусе. Применив специфический ингибитор креатинкиназы (1-фтор-2,4-динитрофенол), а также с помощью агентов, препятствующих окислительному фосфорилированию АДФ в АТФ, Т. Кейн и соавт. (1962) смогли продемонстрировать прямой распад АТФ с одновременным приростом неорганического фосфата и АДФ при одиночном сокращении изолированной мышцы лягушки. Некоторое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции:
Запасы креатинфосфата в мышце невелики, а доступность энергии креатинфосфата имеет ценность для работающей мышцы только в том случае, если расход его постоянно возмещается синтезом АТФ в процессе метаболизма. Для любой ткани, в том числе мышечной, известны два фундаментальных биохимических процесса, в ходе которых регенерируются богатые энергией фосфорные соединения. Один из этих процессов – гликолиз, другой – окислительное фосфорилирование. Наиболее важным и эффективным из них является последний. При достаточном снабжении кислородом мышца, несмотря на анаэробный механизм сокращения, в конечном итоге работает за счет энергии, образующейся при окислении (в цикле Кребса) как продуктов распада углеводов, так и ряда других субстратов тканевого дыхания, в частности жирных кислот, а также ацетата и ацетоацетата. Содержание АТФ и креатинфосфата в сердечной мышце ниже, чем в скелетной мускулатуре, а расход АТФ велик. В связи с этим ресинтез АТФ в миокарде должен происходить намного интенсивнее, чем в скелетной мускулатуре. Для сердечной мышцы теплокровных животных и человека основным путем образования богатых энергией фосфорных соединений является путь окислительного фосфорилирования, связанный с поглощением кислорода. Регенерация АТФ в процессе анаэробного расщепления углеводов (гликолиз) в сердце человека практического значения не имеет. Именно поэтому сердечная мышца очень чувствительна к недостатку кислорода. Характерной особенностью обмена веществ в сердечной мышце по сравнению со скелетной является также то, что аэробное окисление веществ неуглеводной природы при работе сердечной мышцы имеет большее значение, чем при сокращении скелетной мышцы. Только 30–35% кислорода, поглощаемого сердцем в норме, расходуется на окисление углеводов и продуктов их превращения. Главным субстратом дыхания в сердечной мышце являются жирные кислоты. Окисление неуглеводных веществ обеспечивает около 65–70% потребности миокарда в энергии. Из свободных жирных кислот в сердечной мышце особенно легко подвергается окислению олеиновая кислота.
Биохимические изменения при мышечных дистрофиях и денервации мышц. Креатинурия. Общими для большинства заболеваний мышц (прогрессирующие мышечные дистрофии, атрофия мышц в результате их денервации, тенотомия, полимиозит, некоторые авитаминозы и т.д.) являются резкое снижение в мышцах содержания миофибриллярных белков, возрастание концентрации белков стромы и некоторых саркоплазматических белков, в том числе миоальбумина. Наряду с изменениями фракционного состава мышечных белков при поражениях мышц наблюдается снижение уровня АТФи креатинфосфата. Например, через 12 дней после денервации содержание АТФ в денервированной икроножной мышце кролика снижается более чем в 2 раза. Отмечаются также снижение АТФазной активности контрактиль-ныхбелков(миозина), уменьшение количества имидазолсодержащих дипептидов. При прогрессирующих мышечных дистрофиях и других заболеваниях, связанных с распадом мышечной ткани, часто отмечаются сдвиги в фосфолипидном составе мышц: значительно снижается уровень фосфатидилхолина и фосфатидилэтаноламина,концентрация с фингомиелинаи лизо-фосфатидилхолина повышается. Досихпор истинные механизмы изменения фосфолипидного состава мышечной ткани при патологии не выяснены, неизвестна также роль этих сдвигов в патогенезе мышечных дистрофий. Для многих форм патологии мышечной ткани характерны нарушение метаболиз макреатина и его усиленное выделение смочой (креатинурия). Несмотря на многочисленные исследования и обилие фактического материала, вопрос о причинах креатинурии при заболеваниях мышц не может считаться окончательно решенным. Принято считать, что креатинурия у больных миопатией является результатом нарушения в скелетной мускулатуре процессов фиксации (удержания) креатина и его фосфорилирования. Если нарушен процесс синтез акреатинфосфата, то не образуется и креатинина; содержание последнего в моче резко снижается. В результате креатинурии и нарушения синтеза креатинина резко повышается креатиновый показатель (креатин/креатинин)мочи. При патологии мышечной ткани можно наблюдать определенную закономерность в изменении активности ферментов в мышцах: уменьшается активность ферментов, локализованных в саркоплазме; незначительно изменяется активность ферментов, связанных с митохондриями; заметно возрастает активность лизосомальных ферментов. Наконец, показано, что при многих заболеваниях мышечной системы наступают сдвиги в системе цАМФ: снижается содержание цАМФ в мышечной ткани, повышается активность фосфодиэстеразыи нарушается способность аденилатциклазы активироваться под влиянием адреналина и фторида натрия.
Химический состав нервной ткани. Энергетический обмен в нервной ткани. Биохимия возникновения и проведение нервного импульса. Медиаторы: ацетилхолин, катехоламины, серотонин, гамма-аминомасляная кислота, глутаминовая кислота, глицин, гистамин.
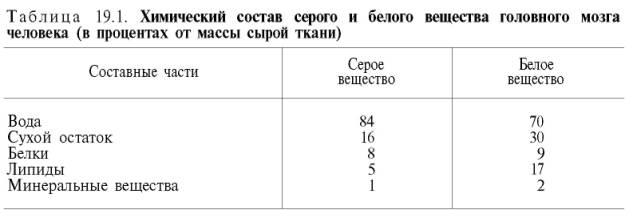
На долю белков приходится примерно 40% от сухой массы головного мозга. Мозговая ткань является трудным объектом для изучения белкового состава вследствие большого содержания липидов и наличия белково-липидных комплексов. А.Я. Данилевский впервые разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки. Обширные исследования в этой области были проведены также А.В. Палладиным и сотр., которые разделили белки нервной тканина 4 фракции: извлекаемые водой, 4,5%растворомКСl, 0,1%растворомNaOH и нерастворимый остаток. Установлено, что серое вещество богаче белками, растворимыми вводе, чем белое вещество,– соответственно 30 и 19%. Белое вещество, напротив, содержит гораздо больше (22%) нерастворимого белкового остатка, чем серое вещество(5%). В дальнейшем было выделено 5–10 фракций растворимых белков мозга, различающихся по своей электрофоретической подвижности. Энергетический обмен в нервной ткани. На долю головного мозга приходится 2–3% от массы тела. В то же время потребление кислорода головным мозгом в состоянии физического покоя достигает 20–25% от общего потребления его все морганизмом, а у детей в возрасте до 4 лет мозг потребляет даже 50%кислорода, утилизируемого все морганизмом. О размерах потребления головным мозгом из крови различных веществ, в том числе кислорода, можно судить по артериовенозной разнице. Установлено, что во время прохождения через мозг кровь теряет около 8 об.% кислорода. В 1 мин на 100 г мозговой ткани приходится 53–54 мл крови. Следовательно, 100 г мозга потребляет в 1 мин 3,7 мл кислорода, а весь головной мозг (1500 г) – 55,5 мл кислорода. Газообмен мозга значительно выше, чем газообмен других тканей, в частности он превышает газообмен мышечной ткани почти в 20 раз. Интенсивность дыхания для различных областей головного мозга неодинакова. Например, интенсивность дыхания белого вещества в 2 раза ниже, чем серого(правда, в белом веществе меньше клеток). Особенно интенсивно расходуют кислород клетки коры мозга и мозжечка. Поглощение кислорода головным мозгом значительно меньше при наркозе. Напротив, интенсивность дыхания мозга возрастает при увеличении функциональной активности.
Биохимия возникновения и проведения нервного импульса. Молекулярные механизмы синаптической передачи Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов К+ и Na+по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов К+ и значительно меньшей для ионов Na+. Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса. При определенных условиях резко повышается проницаемость мембраны для ионов Na+. В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na+, выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К+. В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраныэлектрически возбудимы. При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na+(примерно в 500 раз) и остается без изменения для ионов К+. В результате ионыNa+устремляются внутрь клетки. Компенсирующий поток ионов К+, направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраныприобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К+ над поступлениемионов Na+– мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na+, вошедшие в нейрон при возбуждении, заменяются на ионы К+. Этот переход происходит против градиентаконцентрации, так как ионов Na+во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na+против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калияи натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Заметим, что миелиновые мембраны, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят вклетку и выходят из нее только в тех местах, где изолятор отсутствует. Как уже отмечалось, миелиновая мембрана состоит из фос-фолипидов, в частности из сфингомиелина, холестерина, а также белков и гликосфинголипидов. Некоторые заболевания, например рассеянный склероз, характеризуются демиелинизацией и нарушением проведения нервного импульса. Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффекторного органа.
Медиаторы: ацетилхолин, катехоламины, серотонин, γ-аминомаслянная кислота, глутаминовая кислота, глицин, гистамин. Ацетилхолин является химическим передатчиком (медиатором) нервного возбуждения; окончания нервных волокон, для которых он служит медиатором, называются холинергическими, а рецепторы, взаимодействующие с ним, называют холинорецепторами. Холинорецептор (по современной зарубежной терминологии — «холиноцептор») является сложной белковой макромолекулой(нуклеопротеидом), локализованной на внешней стороне постсинаптической мембраны. При этом холинорецептор постганглионарных холинергических нервов (сердца, гладких мышц, желез) обозначают какм-холинорецепторы(мускариночувствительные), а расположенные в области ганглионарных синапсов и в соматических нервномышечных синапсах — какн-холинорецепторы(никотиночувствительнные). Такое деление связано с особенностями реакций, возникающих при взаимодействии ацетилхолина с этими биохимическими системами: мускариноподобных в первом случае и никотиноподобных — во втором; м- и н-холинорецепторы находятся также в разных отделахЦНС.
Катехоламины — физиологически активные вещества, выполняющие роль химических посредников и «управляющих» молекул (медиаторов и нейрогормонов) в межклеточных взаимодействиях у животных и человека, в том числе в их мозге; производныепирокатехина. К катехоламинам относятся, в частности, такие нейромедиаторы, какадреналин,норадреналин,дофамин(допамин). γ-Аминомасляная кислота выполняет в организме функцию ингибирующего медиатора центральной нервной системы. При выбросе ГАМК всинаптическую щельпроисходит активация ионных каналовГАМКA- иГАМКC-рецепторов, приводящая к ингибированию нервного импульса. становлено, что ГАМК является основным нейромедиатором, участвующим в процессах центрального торможения. Под влиянием ГАМК активируются также энергетические процессы мозга, повышается дыхательная активность тканей, улучшается утилизация мозгом глюкозы, улучшается кровоснабжение. Действие ГАМК в ЦНС осуществляется путём её взаимодействия со специфическими ГАМКергическими рецепторам Глутаминовая кислота является нейромедиаторной аминокислотой, одним из важных представителей класса «возбуждающих аминокислот». Связывание аниона глутамината со специфическими рецептораминейроновприводит к возбуждению нейронов. Существуют ионотропные иметаботропные(mGLuR 1-8) глутаминатные рецепторы. Ионотропными рецепторами являются NMDA-рецепторы,AMPA-рецепторыикаинатные рецепторы. Эндогенные лиганды глутаминатных рецепторов — глутаминовая кислота и аспарагиновая кислота. Для активации НМДА рецепторов также необходим глицин. Блокаторами NMDA-рецепторовявляютсяPCP,кетамин, и другие вещества. AMPA-рецепторы также блокируются CNQX,NBQX.Каинова кислотаявляется активатором каинатных рецепторов. Глицин является нейромедиаторной аминокислотой. Рецепторы к глицину имеются во многих участкахголовного мозгаиспинного мозгаи оказывают «тормозное» воздействие нанейроны, уменьшают выделение из нейронов «возбуждающих» аминокислот, таких, какглутаминовая кислота, и повышают выделениеГАМК. Серотонин играет рольнейромедиатора в ЦНС. Серотонинергические нейроны группируются встволе мозга: вваролиевом мостуи ядрах шва. От моста идут нисходящие проекции вспинной мозг, нейроны ядер шва дают восходящие проекции к мозжечку, лимбической системе, базальным ганглиям, коре. При этом нейроны дорсального и медиальногоядер швадают аксоны, различающиеся морфологически, электрофизиологически, мишенями иннервации и чувствительностью к некоторым нейротоксическим агентам, например,экстази.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-05-27; просмотров: 156; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.27.244 (0.012 с.) |