Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Непосредственное сравнение геномов
Сравнения между ВИЧ и другими вирусами проводились с момента публикации первых последовательностей геномов ретровирусов. Одной из первых работ мы обязаны Гонде и др. (1986). Это очень уважаемая исследовательская группа, в которой на этот раз также участвовал Роберт Галло.
Прямое сравнение геномов ВИЧ и вируса Висна показало, что 60 процентов нуклеотидов идентичны. Это было гораздо лучшее совпадение, чем между ВИЧ и любым другим ретровирусом. Это соответствие последовательностей особенно очевидно проявилось в работе МакКлюра. Последовательности для кодирования трех разных белков были использованы для построения генеалогического древа ряда различных ретровирусов. Рис. 13 на следующей странице показывает, что во всех трех случаях рассматриваемый ген ВИЧ имеет общее происхождение, то есть либо вирус Visna происходит от ВИЧ, либо ВИЧ происходит непосредственно от вируса Visna. Но поскольку вирус Visna был известен задолго до возникновения СПИДа, мы можем только сделать вывод, что ВИЧ является ответвлением вируса Visna. Никакой другой известный ретровирус для этого не подходит.
Рис 13. Родословная различных ретровирусов, установленная путем компьютерного сравнения геномов трех разных белков: обратной транскриптазы, рибонуклеазы H и белка оболочки. Маркировка стрелкой добавлена автором. Обратите внимание, что во всех трех случаях ВИЧ ответвляется непосредственно от вируса Visna. (МакКлюр и др.)
Для постороннего совпадение 60% может быть разочаровывающим, но это именно то, чего и следовало ожидать. Вирусы, как известно, очень изменчивы, но ВИЧ превосходит их все. Он мутирует примерно в миллион раз чаще, чем, например, вирус Visna. Обычно считается, что это происходит из-за ненадежной обратной транскриптазы.
Хахн в лаборатории Галло каждые два года изолировал ВИЧ от одного и того же больного СПИДом. Они обнаружили, что вирус изменился примерно на 10 процентов за два года. Весной 1986 года Гонда провел свой эксперимент. Весной 1984 года разница между вирусом Визна и ВИЧ должна была составлять не 40 процентов, а всего 30 процентов, весной 1982 года - только 20 процентов, весной 1980 года - не более 10 процентов, а весной 1978 года - ноль процентов. Это означало бы, что весной 1978 г. ВИЧ отделился от вируса Висна и начал свою независимую эволюцию в организме пациентов. Этот «день рождения» вируса СПИДа также соответствует тому факту, что первые случаи СПИДа произошли в Нью-Йорке летом 1979 года.
В этой связи мы должны иметь в виду, что в случае полового заражения инкубация занимает два года или даже намного дольше до начала полноэкранного СПИДа, но в случае массивных инфекций, например, при переливании крови, инкубация часто наблюдаются времена от одного до полутора лет. Таким образом, первые случаи полноэкранного СПИДа летом 1979 года указывают на то, что вирус был завершен в начале 1978 года, что в точности совпадает с результатом расчета регрессии, основанного на частоте мутаций.
Чтобы понять высокую скорость мутаций в ретровирусах, мы должны рассмотреть механизм обратной транскрипции. Цепь РНК из более чем 9000 нуклеотидов образует спираль из примерно 900 витков. Если бы нужно было скопировать спираль ДНК одинаковой длины в связную цепь, они никогда бы не разошлись. Следовательно, обратная транскрипция происходит на коротких участках ДНК, состоящих примерно из полутора витков спирали, которые легко отделяются от матрицы РНК. Вторая цепь ДНК реплицируется на этих коротких цепях ДНК, и только тогда эти короткие двойные цепи склеиваются вместе, образуя когерентную цепь ферментной лигазой. Между тем, они подвержены броуновскому движению молекул, они могут выпрыгивать из линии, перемещаться не в то место или просто развернуться. Соответственно, мутации ретровирусов предпочтительно относятся к типу делеции (сбой), вставки или инверсии. Чем меньше времени проходит до того, как фрагменты свариваются лигазой, тем меньше мутаций. Следовательно, различные ферменты, участвующие в процессе, должны работать оптимально быстро и точно согласовывать свое действие. Но даже в этом случае невозможно полностью избежать неправильных копий и мутаций.
Но что произойдет, если вирус будет собран с помощью генной инженерии из двух частей генома, которые не совсем соответствуют друг другу? Ферменты не работают оптимально, фрагменты ДНК медленно образуются и медленно слипаются. Это должно было быть особенно плохо, если обратная транскриптаза была включена в геном вируса Visna другим вирусом. Это действительно так.
Chandra et al. изолировал обратную транскриптазу от возбудителя СПИДа ВИЧ. Оказалось, что этот фермент состоит из двух отдельных фракций, которые также имеют оптимальную функцию при различных значениях pH, один раз при pH 6,3, а другой - при pH 5,8, то есть с двумя разными кислотными значениями (Рисунок 15). По нашей просьбе Чандра сообщил нам, что вирус Visna обладает одной из этих фракций, HTLV-I - другой, и что обе эти фракции вместе могут быть обнаружены только в ВИЧ. Он сказал, что это также является причиной неточной обратной транскрипции и частых мутаций ВИЧ. Все ведет себя так, как если бы небольшой фрагмент генома HTLV-I был вставлен в вирус Visna, который кодирует часть обратной транскриптазы в этом вирусе.
Рис. 15. Активность обратной транскриптазы ВИЧ как функция pH. (Чандра и др.)
Сравнивая геномы ВИЧ и Visna, Коффин (1986) действительно обнаружил, что геном ВИЧ был примерно на 300 нуклеотидов длиннее, чем геном Visna. Это соответствует кодированию 100 аминокислот, которых было бы достаточно для белкового мономера, субмолекулы, которой было бы достаточно, чтобы нести активный центр молекулы фермента. Все ведет себя так, как будто часть генома HTLV-I с кодированием обратной транскриптазы была дополнительно вставлена в (полный) геном Visna.
Таким образом, вирус СПИДа будет рекомбинантной конструкцией вируса типа C и вируса типа D с двумя ферментными системами, которые не скоординированы друг с другом. В этих условиях такая деликатная операция, как обратная транскрипция, не может пройти без множества ошибок. Работа Чандры дает нам прямые доказательства рекомбинантной природы ВИЧ.
Сравнение генома методом гетеродуплексной гибридизации Прежде чем мы сообщим о впечатляющих открытиях, которые предлагает нам этот метод, мы должны сначала сделать несколько пояснений. Известно, что двухцепочечная ДНК состоит из двух комплементарных нуклеотидных цепей, которые соединяются друг с другом посредством многочисленных водородных связей. Если разорвать их на части, они вскоре снова сойдутся в правильном порядке, они гибридизуются. Если две такие цепи принадлежат к двум разным типам, они будут гибридизоваться только при наличии хотя бы некоторой степени сходства в последовательностях. Степень гибридизации дает нам информацию о степени родства между двумя геномами.
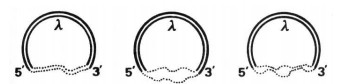
Гибридизация гетеродуплексов также дает нам картину распределения участков генома, которые гибридизуются друг с другом. Для этой цели кольцевой двухцепочечный геном ДНК, например геном бактериофага λ (греческая буква лямбда; здесь обозначение типа бактериофага), разрезается и вставляются две нити протовируса ДНК или две одиночные нити. в разрыв двух разных протовирусов, которые нужно сравнить. Наконец, ионная сила окружающего раствора регулируется так, чтобы цепи ДНК в достаточной степени прилипали друг к другу, если они подходят на 40, 60 или 80 процентов. В случае, который мы собираемся обсудить, была выбрана строгость в 60 процентов, что означает, что две цепочки будут держаться вместе, если они совпадут на 60 процентов или более.
На Рис.16 схематично показан этот способ реакции. На фиг.16а две цепи ДНК согласованы друг с другом по всей своей длине и, следовательно, образуют толстую двойную цепь. На фиг.16b они совершенно не соответствуют друг другу, не гибридизуются, а, скорее, остаются двумя отдельными тонкими одиночными цепями. На фиг.16c их структура соответствует в двух областях. Там они образуют двойные нити, а между ними остаются открытые петли. Такие фигуры можно сфотографировать в электронный микроскоп и оценить графически, как показано на Рис.
Рис. 16 Механизм гибридизации гетеродуплекса
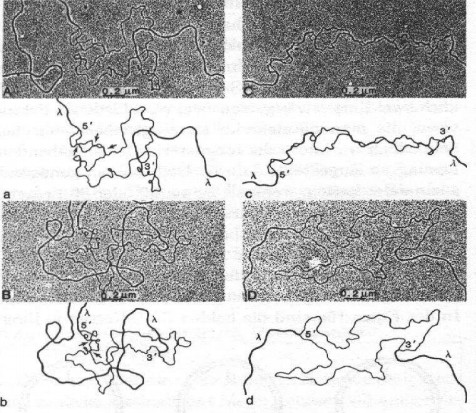
Рис 17 Гибридизация гетеродуплекса между геномами разных ретровирусов (Gonda et al., 1985) Вверху: изображение электронного микроскопа Ниже: упрощенная графическая транскрипция В а) и б) стрелки указывают области гибридизации. А и А: HTLV-I и ВИЧ; B и b: HTLV-I и Visna; C и c: ВИЧ и VisnaVirus; D и d: ВИЧ и инвертированный вируса Висны
Рис 17 был получен лабораторией Гонды (Гонда, 1985). https://pubmed.ncbi.nlm.nih.gov/2990686/ https://pubmed.ncbi.nlm.nih.gov/2981428/
Рисунки A, B, C и D представляют собой записи, сделанные с помощью электронного микроскопа, а расположенные под ними рисунки a, b, c и d представляют собой соответствующие им упрощенные графические изображения.
На Рис.17 B и b мы видим гетеродуплексную гибридизацию между HTLV-I и вирусом Visna. Они гибридизуются только в двух узких областях, обозначенных стрелками. Это никоим образом не говорит о тесной связи между двумя вирусами. Эти два района соответствуют области gag (ген, связанный с геномом); они кодируют белки, которые вместе с РНК образуют ядро, внутреннее тело, которое играет роль в вирусах, аналогичную роли ядра в высших клетках. Структура этих белков адаптирована к взаимодействию с РНК и поэтому имеет практически идентичные участки во всех ретровирусах, что также отражается на структуре генома. Таким образом, гибридизация в диапазоне кляпов не свидетельствует о тесной связи.
На Рис.17 C и c мы видим гетеродуплекс между вирусом Visna и ВИЧ. Два генома гибридизуются на больших расстояниях, что означает, что они совпадают по более чем 60 процентам нуклеотидов. В остальных местах остаются открытые петли; там согласие меньше 60 процентов. В целом, эти два генома гибридизуются в 25 местах, что предполагает тесную связь между двумя вирусами. Согласование более 60 процентов соответствует значениям, которые были позже определены путем сравнения двух последовательностей генома с компьютером.
Чтобы проверить, в какой степени случайные совпадения могут моделировать тесную взаимосвязь, две нити ДНК ВИЧ и вируса Visna были включены в геном-носитель в противоположных направлениях на фиг.17D и d. Никакой гибридизации нет. Если на других фигурах цепи гибридизируются друг с другом, это говорит о реальной взаимосвязи между двумя вирусами, используемыми в каждом случае.
Рисунки 17A и a предоставляют нам наиболее интересную информацию. Они показывают гетеродуплекс между ВИЧ и HTLV-I. Конечно, они гибридизуются в двух узких областях, которые мы уже определили между вирусом Visna и HTLV-I, но есть также область шириной около 300 нуклеотидов. Основываясь на работе Коффина, мы придерживаемся предположения, что сегмент из 300 нуклеотидов из генома HTLV-I был вставлен в геном вируса Visna. Здесь у нас есть фотография этого раздела.
Совершенно невозможно, чтобы такой участок мог возникнуть спонтанно из-за случайных мутаций в геноме Вишны. Мы подсчитали вероятность того, что серия совпадений могла бы привести даже к пятидесятипроцентной идентичности на участке длиной 300 нуклеотидов. Эта вероятность составляет 1: 6 x 10 -82, так что она бесконечно мала и совершенно не может рассматриваться как причина. Нечто подобное может произойти только при целевой передаче. В дополнение к открытиям Чандра, Рис.17A и a также доказывают, что ВИЧ является продуктом генетической рекомбинации вируса Visna и HTLV-I.
В каждом геноме ретровируса есть раздел под названием pol, pol - это сокращение от полимеразы. В этом разделе содержатся гены различных ферментов, которые строят цепи ДНК и РНК из отдельных нуклеотидов. Все они известны как полимеразы. Среди них обратные транскриптазы играют уже описанную важную роль. Только что упомянутый диапазон из 300 нуклеотидов, который, очевидно, происходит от HTLV-I, находится в сегменте полимераз. Она вполне может соответствовать второй обратной транскриптазе ВИЧ, которая, как продемонстрировал Чандра на Рис.15, происходит от HTLV-I.
Конструирование ВИЧ Если наше предположение верно, то вирус СПИДа был создан путем добавления раздела из HTLV-I в геном вируса Висна, который вызывает болезнь, которая смертельна для овец, но не поражает людей, позволив вирусу прикрепляться к человеческим Т4-клеткам. Известно, что ВИЧ, как и большинство других вирусов, может паразитировать на любой клетке, только если ему удастся проникнуть через клеточную мембрану внутрь клетки. С помощью искусственной трансфекции клеток мышечной или соединительной ткани человека или мышей, передаваемый ВИЧ, без каких-либо проблем размножился там. Вирус овцы, которому было предоставлено дополнительное устройство для присоединения к человеческим Т4-клеткам и поглощения ими, должен был проявить свой патогенный эффект и у людей и стать смертельным агентом для них.
Нас часто обвиняли в том, что мы отложили создание ВИЧ до 1977 года. В то время генная инженерия была недостаточно развита, чтобы успешно провести такую сложную операцию. Мы должны этому категорически возразить. В 1977 году в США уже было восемь коммерческих компаний, успешно занимавшихся генной инженерией. Конечно, целенаправленная трансплантация точно определенного участка генома в точно определенное место другого генома - сегодня это работа нескольких дней для технического ассистента в специальной лаборатории - тогда была невозможна. Но уже было известно около 50 рестрикционных ферментов, то есть ферментов, которые разрезают нуклеотидную цепь в точно определенной точке. Уже было известно, что лигазы склеивают разрезанные цепи ДНК. Очень сложный гетеродуплексный метод, при котором вирусные геномы имплантируются в желаемой ориентации в геномы бактериофагов, был известен с 1971 года.
На практике в то время применялся так называемый метод дробовика. В случае конструирования ВИЧ это выглядело бы так. Геномы вируса Visna изначально были включены в геномы бактериофагов, как и гетеродуплекс. Затем они были разрезаны рестрикционным ферментом в одной точке. В другом сосуде геномы HTLV-I были разрезаны на множество мелких фрагментов несколькими рестрикционными ферментами, и эта смесь была добавлена к разрезанным геномам Visna. Затем пробелы в геномах Visna были снова закрыты лигазами с включением того или иного участка генома HTLV-I, в зависимости от случая. Из множества рекомбинантных геномов, полученных таким образом, те, которые проявляли желаемые свойства, были выделены путем отбора и затем размножены. Например, если вам нужны были новые вирусы, которые могли бы воспроизводиться в клетках Т4 человека, вам нужно было только начать культивирование с клетками Т4. Размножаться могли только вирусы, прикрепленные к клеткам Т4. Остальные были ликвидированы.
Среди бесчисленных комбинаций, которые возникли в результате процесса дробовика, неизбежно должны были возникнуть вирусы, которые в дополнение к нормальному гену обратной транскриптазы, несущемуся в каждом геноме Visna, получили второй такой ген от HTLV-I. В результате получился вирус с двумя обратными транскриптазами с двумя функциональными оптимумами, которые мешали друг другу и приводили к бесчисленным мутациям. Коффин, Хан и др. обнаружили, что среди различных белков ВИЧ белок gp120, с помощью которого вирус прикрепляется к клетке Т4, особенно подвержен мутациям. Небольшая часть gp120, которая используется непосредственно для прикрепления вируса, особенно подвержена мутации. С многочисленными мутациями, которые были вызваны двумя разными обратными транскриптазами, неизбежно также должны были возникнуть варианты, которые позволяли прикрепляться к клетке Т4 и которые могли быть выделены и размножены путем отбора. Вирусы, которые наиболее надежно прикреплены к клетке Т4, будут размножаться, и создание и отбор ВИЧ на средства 1977 года будут завершены. Выполнение работы, включая довольно длительный отбор, должно было занять тогда около шести месяцев."
В более ранних публикациях мы представляли этот процесс в упрощенном виде и предполагали, что часть гена из HTLV-I со связывающим эпитопом для рецептора CD4 человеческих Т4-клеток была перенесена непосредственно в геном вируса Visna. Работа Чандры над двумя отдельными оптимумами активности обратной транскриптазы, а также анализ геномных последовательностей современными компьютерными методами, которые мы сразу обсудим, показали нам, что эта гипотеза была слишком простой и позволила нам более правильно представить конструкцию ВИЧ..
Вопреки многим заявлениям в прессе, наша презентация полностью соответствует последним экспериментальным данным. Эти результаты частично основаны на гетеродуплексном методе, частично на компьютерном сравнении нуклеотидных последовательностей разных геномов. Гетеродуплекс - полностью объективный метод. Если он говорит, что существует среднее 60-процентное сходство между вирусом Висна и ВИЧ, но не между HTLV-I и ВИЧ, то это неопровержимый факт. И если этот метод говорит, что HTLV-I и ВИЧ в диапазоне pol имеют на 60 процентов идентичную группу из примерно 300 нуклеотидов, то это соответствует объективной реальности, с которой должна бороться каждая теория.
Чтобы избавиться от этого камня преткновения, было сделано много попыток изобразить гетеродуплекс как немодный и неточный процесс. Это ужасное искажение. Конечно, в отличие от компьютерного сравнения, оно не может сказать, идентична ли определенная пара нуклеотидов в двух вирусах или нет, но его утверждения абсолютно объективны и не могут быть подавлены или подчеркнуты компьютерной программой, выбранной автором. Если гетеродуплекс указывает на отсутствие какой-либо идентичности между геномами HTLV-I и ВИЧ более 90 процентов их длины, то это означает, что ВИЧ ни при каких обстоятельствах не может развиться спонтанно из вируса группы HTLV / SIV; Таким образом, ни от HTLV-I, ни от одного из многочисленных вирусов обезьян, которые мы теперь имеем под собирательным термином SIV (обезьяний вирус иммунодефицита = вирус иммунодефицита обезьян). И если есть совпадающий участок в 300 нуклеотидов между HTLV-I и ВИЧ, который не может быть основан на совпадении, необходимо объяснить, как этот чужеродный участок попал в геном ВИЧ. Наша теория о том, что часть домена pol был генетически преобразована в геном вируса Visna, объясняет оба факта.
Никакая другая теория даже не пыталась объяснить это. Авторы всегда говорят, что ВИЧ естественным образом развился из другого вируса, но никто еще не хотел описать нам этот естественный путь, и никто не смог назвать вирус, из которого мог развиться ВИЧ.
В естествознании есть испытанный принцип. Явление должно быть объяснено известными структурами, функциями и законами природы. Если не существует другой теории, основанной на естественных элементах, единственная существующая должна быть признана научно обоснованной на данный момент. Ссылки на «любой еще не обнаруженный вирус» или на «неизвестный нам путь» принципиально отвергаются. Если возникновение или возникновение ВИЧ было чисто научной проблемой без какой-либо политической подоплеки, наша теория должна быть пока единственной.
Конечно, мы также должны требовать от проверенной научной теории, чтобы она не противоречила подтвержденным экспериментальным результатам. Сейчас была сделана попытка продемонстрировать такое противоречие между результатами гетеродуплекса, на которых мы основываем наши аргументы, и компьютерным сравнением геномов. Однако этот аргумент не выдерживает более внимательного изучения.
Давайте сравним, например, площадь pol ВИЧ с областью pol вируса Visna и HTLV-I в соответствии с методами, описанными Куртом опубликовавшим компьютерные сравнения. В ВИЧ pol состоит из 3074 пар нуклеотидов. 300 из них были заимствованы из HTLV-I, оставшиеся примерно 2770 пар нуклеотидов должны, если Гонда правильно измерил, образовать несколько групп, более половины из которых имеют идентичность 60 или более процентов. Фактически, компьютер находит три группы из 757, 64 и 766 пар нуклеотидов в области pol, в которых идентичность между ВИЧ и вирусом Visna составляет от 58 до 62 процентов, то есть для 1583 пар нуклеотидов, то есть более половины в области pol эти два вируса совпадают точно так же, как Гонда обнаружил гетеродуплексным методом.
Другим важным моментом является наличие 60-процентного идентичного участка из 300 пар в области pol ВИЧ и HTLV-I. Компьютерное сравнение показывает нечто подобное, но только для диапазона из 132 пар нуклеотидов с идентичностью 58 процентов. Но ничего лучше ожидать не приходилось. Гонда сравнил геном Visna с геномом изолята ВИЧ из начала 1985 года и получил 60-процентную идентичность для секции из 300 пар. Курт и др. сравните его с изолятом 1988 года, который, как ожидается, мутировал примерно на 15 процентов по сравнению с изолятом 1985 года. С чисто статистической точки зрения идентичность 58% могла быть сохранена только в части всего раздела. Здесь мы также находим полное подтверждение результатов гетеродуплекса. < Аргумент против нашей теории ни в коем случае не выводится из значений полученных Куртом? >.
Собственно говоря, эта теория исходит от нас лишь в части деталей. Предположение, что вирус СПИДа является результатом генно-инженерной комбинации двух ранее известных вирусов, было сформулировано еще в 1984 году на конференции AAAS, но не было зафиксировано в протоколе. Заявление Алисона и Монтанье, сделанное в том же году, что HTLV-I не мог быть преобразован, попадает в ту же ноту. Потому что тогда остается только один искусственный путь - генная инженерия рекомбинантного вируса.
В следующей главе мы рассмотрим некоторые политические аспекты происхождения СПИДа. Аргументы в этой области имеют совсем другой характер. Здесь нет никаких экспериментальных доказательств. Мы полагаемся на документы, и большая их часть до сих пор держится в секрете. Но даже те немногие, которые были обнародованы, с полной уверенностью подтверждают, что возбудитель СПИДа был намеренно произведен в военной лаборатории США. Таким образом, наша теория, основанная на подробных биологических анализах, подтверждается официальными документами совершенно иным образом." [1 стр 57-68]
"Наш тезис, согласно которому вирус СПИДа был получен в американской военной лаборатории с использованием процессов генной инженерии, вызвал многочисленные отклики. Большинство из них пытаются произвести впечатление на неподготовленную аудиторию, и их легко опровергнуть. Но они получили широкое распространение через СМИ. Поэтому мы обязаны рассмотреть их здесь. Чаще всего мы слышали аргумент о том, что СПИД - это медленно развивающееся заболевание, и поэтому вирус СПИДа не мог быть создан с намерением использовать его в качестве оружия войны. Мы уже рассматривали этот вопрос в Разделе 8.4 и хотим лишь кратко резюмировать здесь ответ. Если вам нужен новый вирус, который нельзя обнаружить с помощью антител, вам нужно начать с ретровируса, геном которого транскрибируется с одноцепочечной РНК в двухцепочечную ДНК. Это не имеет антигенного действия и не обнаруживается антителами, направленными против компонентов вируса. В 1977 году был известен только один патогенный и смертельный ретровирус, вирус Висна, а также вирус, который атакует клетки Т4 человека и в основном не является патогенным, который позже был описан как HTLV-I. Результатом их рекомбинации стал ВИЧ, смертельный патоген для людей, но не удачный в качестве оружия войны.
Сегодня мы знаем, что вирус Visna - это «медленный вирус». На его основе вряд ли можно создать биологическое оружие, которое убивает быстро и надежно. Но про 1977 год никто не знал. Обычно считалось, что болезнь Висны прогрессирует медленно, поскольку в Исландии овец содержали на открытом воздухе и в здоровой окружающей среде. Ведь больной туберкулезом долго выживает в чистом горном воздухе, а в городской квартире с тусклым светом быстро умирает. Сегодня мы знаем, что рекомбинантный вирус HTLV-I / Visna также остался лентивирусом, но кто мог предположить это?
РОБЕРТ СТЕККЕР: быстрые вирусы, конечно, бесполезны, потому что от них слишком легко защититься. Я имею в виду, что очень быстро действующий вирус никуда не годится. ЛЕОНАРД ГОРВИЧ: Что вы имеете ввиду в виду? РОБЕРТ: Фрэнк Феннер говорит обо всех характеристиках…<..>., Колд-Спрингс-Харбор предложили на конференции грандиозную вещь, модель MMMV «maximally monstrous malignant virus» («максимально чудовищный злокачественный вирус»), а затем дали все характеристики, которыми он должен обладать. И они говорили о том, что нужно, чтобы произвести такой вирус. И, конечно же, все характеристики в точности такие же, как у вируса СПИДа, за исключением одного, а именно передачи в виде аэрозоля, что, как мы считаем, потенциально возможно.
[Результат поиска по MMMV: “На конференции в середине 1980-х годов Килборн придумал сценарий о вымышленном вирусе кошмара, обладающем свойствами, которые сделают его самым заразным, самым смертоносным и самым трудным для контроля. Он назвал его “максимально злокачественным (чудовищным) вирусом”, или МММВ. Как описал Килборн, среди других гнусных атрибутов МММВ, он будет передаваться по воздуху, как грипп, будет экологически устойчивым, как полиомиелит, и будет вставлять свои собственные гены непосредственно в ядро хозяина, как ВИЧ.”[ссылка]]
ЛЕН. [Ой, не дай бог, - подумал я [ГОРВИЦ]. Раньше я не слышал этой теории. Учитывая очевидный интеллект и огромные знания Стрекера, его утверждение поразило меня.] РОБЕРТ. Но они подготовили статьи о том, что делает вирусы опасными и чудовищными. И одна из вещей - они работают медленно, а не быстро. И что они постоянно видоизменяются. В точности характеристики СПИДа. ЛЕН: Интересно. Это невероятно. РОБЕРТ. Да, это так.
Профессор Карин Мёллинг из Института Макса Планка в Западном Берлине возражает, что примерно в 1977 году генетических знаний было слишком мало для достижения такого результата. Для этого потребовался бы гений масштаба Роберта Галло. И она спрашивает: «А был ли у Пентагона второй Галло?» Г-жа Меллинг должна знать, что Галло был рядом - в качестве директора онкологического института в Бетесде, штат Мэриленд, а с 1975 года - в качестве главы вирусного отдела в Форт-Детрик., Мэриленд. Он уже изолировал вирус AL23V в 1975 году, а затем попытался скрыть его, опубликовав семь лет спустя под названием HTLV. Нет, Пентагону не нужен был второй Галло. Он купил Галло №1.
Открытие фермента обратной транскриптазы было опубликовано в 1970 году. Но за много лет до этого Темин и Балтимор уже исследовали и описали обратную транскрипцию, столь характерную для ретровирусов. Еще в 1969 году Пентагон смог объявить, что должна быть возможность создать патогенный вирус, который ускользнет от иммунной системы инфицированного организма. Выдающиеся ученые заверили, что это возможно и технически (см. Приложение I).
Профессор Кох из Института Роберта Коха в Западном Берлине и д-р. Лёвер, заместитель директора Института Пауля Эрлиха во Франкфурте-на-Майне, утверждал, что в 1977 году генная инженерия была недостаточно развита, чтобы можно было сконструировать ВИЧ. Здесь мы должны категорически не согласиться. В Разделе 6.2.3 мы описали гетеродуплексный метод, который используется Гондой для гибридизации геномов. Что касается технологии, они ссылаются на публикацию 1971 года, что означает, что самое позднее к 1970 году основные элементы этой чрезвычайно сложной генетической манипуляции уже были доступны. С тех пор были внесены дальнейшие улучшения. Ведь недаром еще в 1978 году возникло не менее восьми коммерческих компаний, основанных на генной инженерии.
Чтобы завершить это обсуждение, мы попытались разработать стратегию, с помощью которой стало бы возможным создание ВИЧ с ресурсами 1977 года. В то время было известно по крайней мере 50 рестрикционных ферментов, то есть ферментов, которые разрезают цепь нуклеиновых кислот в определенном месте - сегодня мы знаем их более 200. Мы также знали лигазы, ферменты, которые соединяют такие границы раздела вместе.
Чтобы собрать ВИЧ, необходимо сначала интегрировать геном вируса Visna в геном-носитель, например, в геном бактериофага, как это делается с помощью гетеродуплексной техники. Геном Visna можно разрезать в одной точке с помощью любого рестрикционного фермента. Геномы HTLV-1 измельчаются на различные фрагменты многочисленными рестрикционными ферментами, и этот «фарш» смешивается с разрезанными геномами вируса Visna. Если сейчас добавить лигазу, то тот или иной фрагмент будет случайно встроен в пробел в геноме ВИЧ. Если эти различные продукты выращивать в культуре клеток Т4 человека, только те, которые могут прикрепляться к клеткам Т4, будут размножаться. Вирусы с желаемыми новыми свойствами можно выбирать из множества продуктов и размножать.
Этот «принцип дробовика» широко использовался в 1970-х годах. Сегодня его вытеснили более точные, целенаправленные методы. Два известных молекулярных биолога подтвердили нам, что предложенная нами стратегия могла привести к появлению ВИЧ в течение примерно шести месяцев. При сегодняшних методах время составит всего несколько дней.
Совершенно неожиданный аргумент был выдвинут против нас профессором Куртом из Института Пауля Эрлиха и профессором Мёллингом. Они утверждают, что геном ВИЧ намного сложнее, чем у любого другого ретровируса, и поэтому никогда не мог быть создан путем рекомбинации геномов других ретровирусов. Профессор Курт даже требует от нас, чтобы мы называли "отца и мать" ВИЧ по имени, как будто мы не ясно заявили из нашего первого сообщения, что ВИЧ представляет собой рекомбинацию генома вируса Висна с небольшой частью генома HTLV-I.
Профессор Мёллинг идет еще дальше. Она утверждает, что только у ВИЧ есть ген tat и что он не может быть вызван какой-либо рекомбинацией. Ген tat, точнее группа генов tat, является трансактиватором, который заставляет вещества, необходимые для вируса, накапливаться примерно в тысячу раз быстрее.
Даже в таком виде критика недопустима. Несомненно, ген tat и сложная структура 3'-конца ретровирусного генома были впервые изучены на ВИЧ, но вывод о том, что в других Лентивирусах нет ничего аналогичного, был чистым выдаванием желаемого за действительное до тех пор, пока соответствующие исследования не были проведены на других Ретровирусах.
Реальность выглядит еще уродливее. Уже в июне 1986 года Гонда опубликовал сравнительное исследование ВИЧ и вируса Висна, о котором мы сообщили в Разделе 6.2.2. Было обнаружено, что оба генома в значительной степени идентичны и что все структурные элементы присутствуют в обоих, за исключением небольшого участка из 300 нуклеотидов, который соответствует участку HTLV-I. Таким образом, все структурные элементы, впервые описанные в ВИЧ, включая группу tat, должны были присутствовать и в вирусе Visna. Статья из такой уважаемой лаборатории, как Ганда, вряд ли могла ускользнуть от внимания специалистов по СПИДу в ФРГ, и когда в ноябре 1986 года они начали размещать свои аргументы в «Spiegel», то есть не в специализированном журнале, то это расценивается как попытка ввести в заблуждение общественное мнение.
То же самое и с предполагаемым африканским происхождением СПИДа. В последнее время никто не осмеливался публично защищать против нас теорию зеленой обезьяны. Но нас несколько раз обвиняли в пренебрежении многими открытиями по обнаружению антител к ВИЧ в сыворотке крови с 1960-х и 1970-х годов. К сожалению, ни один из этих критиков не упомянул об исследовании, проведенном более современными методами за последний год, которое опровергло все предыдущие выводы. Ниже приводится отрывок из отчета, опубликованного в 1987 году в западногерманском журнале Stern, который показывает шокирующе низкий уровень, на котором ведутся дискуссии о происхождении СПИДа.
Репортер Х.-Х. Клар обвиняет Сигалов в том, что они приписывают открытия признанных ученых техническим ошибкам и использованию устаревших и неподходящих методов. «Но все эти тесты были проверены с помощью высоконадежного и селективного метода вестерн-блоттинга. Здесь не может быть ошибки». На этом репортер останавливается. Его утверждение должно было пойти дальше: «... Этот метод опроверг все предыдущие открытия и показал, что не было доказательств наличия антител против СПИДа у жителей Африки до 1980 года». Но писатель вовремя останавливается, и это так. его отчет - не что иное, как грубое заблуждение.
В 1986 и 1987 годах у нас было несколько возможностей соревноваться с ведущими специалистами по СПИДу на публичных собраниях, и все они имели большой успех. За последние два года ни один организатор не смог найти для нас такого партнера по дискуссии. Кажется, что в области биологии нашим аргументам нечего серьезно противопоставить.
В отсутствие научных аргументов было предпринято несколько попыток доказать предполагаемые случаи СПИДа до 1977 года. Поскольку мы утверждаем, что СПИД был искусственно создан в лаборатории Пентагона в 1977/78 году, такие свидетельства явно опровергают наш тезис. Не проходит и года, чтобы в прессе не сообщалось о случае европейца, который после пребывания в Африке заболел болезнью, которая не могла быть ничем иным, как СПИДом. Все эти истории болезней содержат очевидные несоответствия, так что на сегодняшний день ни одна из них не была признана Всемирной организацией здравоохранения окончательной. В качестве примера рассмотрим самый последний из этих случаев.
Средства массовой информации недавно с большим энтузиазмом восприняли объявление Дж. Корбитта, А.С. Бэйли и Дж. Миллиамса, опубликованное в престижном английском медицинском журнале «The Lancet» (том 336, стр. 51, 1990), что умерший в 1959 г. моряк явно был болен СПИДом. В образцах тканей, которые остались бы от пациента, геном ВИЧ-I был достоверно обнаружен недавно разработанным методом размножения генов.
Значит, это моряк, который, скорее всего, был в Африке и мог там заразиться. Фактически, он умер в 1959 году от пневмоцистной инфекции и цитомегаловируса, двух оппортунистических инфекций, характерных для смерти от СПИДа. Но одно это еще не доказательство. Многие тропические патогены, например червь Schistosoma, дизентерийная амеба, возбудитель малярии Plasmodium, лепра и туберкулезная палочка и другие, обладают иммуно-ингибирующим действием. Заражение двумя или даже тремя из них вызывает обширное нарушение иммунной системы с возникновением оппортунистических инфекций, что во всех отношениях соответствует конечной стадии СПИДа. Смерть британского моряка от иммунодефицита сама по себе не свидетельствует о СПИДе. Даже в профессиональном плане он особенно подвержен тропическим болезням.
ри доктора из Манчестера теперь заявляют, что предоставляют доказательства того, что пациент действительно умер от ВИЧ-инфекции. Это требует некоторых технических пояснений.
В больнице был обнаружен парафиновый блок с расплавленным куском ткани пациента. Для исследования под микроскопом ткани обезвоживают спиртом и бензолом, заливают жидким парафином и после застывания разрезают на срезы толщиной в микрометр. Парафин удаляется, срез окрашивается и просматривается. Оставшиеся парафиновые блоки могут храниться неограниченное время без охлаждения, почти не занимают места и поэтому часто хранятся вместе с файлами пациента.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 89; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.138.144 (0.079 с.) |