Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 1. Общие приниципы организации и классификация тканейСтр 1 из 23Следующая ⇒
МОДУЛЬ 1: «ТКАНИ (ОБЩИЕ ПОНЯТИЯ И ТЕРМИНЫ). ЭПИТЕЛИАЛЬНЫЕ ТКАНИ, ЖЕЛЕЗЫ. КРОВЬ И ЛИМФА. КРОВЕТВОРНЫЕ ТКАНИ. ГЕМОПОЭЗ» Тема 1. ОБЩИЕ ПРИНИЦИПЫ ОРГАНИЗАЦИИ И КЛАССИФИКАЦИЯ ТКАНЕЙ ОБЩИЕ ПРИНИЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ КЛАССИФИКАЦИЯ ТКАНЕЙ Организм человека содержит большое разнообразие тканей, которые наиболее часто объединяют (1) в группы по признакам сходства их строения и функций (морфофункциональный принцип) или (2) в типы на основании общности источников их развития (гистогенетический принцип). Морфофункциональная классификация тканей, впервые предложенная в 50-х г.г. XIX столетия немецкими гистологами Ф. Лейдигом и Р. Келликером, получила наибольшее распространение. Она выделяет четыре группы тканей (см. схему): (1) эпителиальные (пограничные); (2) соединительные (ткани внутренней среды); (3) мышечные и (4) нервную (нейральную).
Каждая группа тканей может включать ряд подгрупп. Внутри отдельной ткани выделяют различные клеточные популяции. Последние могут разделяться далее на индивидуальные субпопуляции. 1. Эпителиальные (пограничные) ткани характеризуются сомкнутым расположением клеток, образующих пласты, практическим отсутствием межклеточного вещества, пограничным положением в организме (обычно на границе с внешней средой), полярностью. Их основные функции - барьерная, защитная, секреторная 2. Соединительные (ткани внутренней среды) - обширная группа, объединяющая ряд подгрупп тканей, общим признаком которых служит резкое преобладание межклеточного вещества по объему над клетками. Эти компоненты в различных тканях этой группы существенно различаются по строению, физико-химическим свойствам, количественному соотношению и пространственной организации. Важнейшие функции соединительных тканей - гомеостатическая, опорная, трофическая, защитная. 3. Мышечные ткани обладают сократительной способностью, благодаря которой они выполняют свою основную функцию - перемещение организма или его частей в пространстве. Морфологически мышечные ткани представлены удлиненными сократимыми элементами (клетками или волокнами), которые обычно располагаются параллельно друг другу и объединены в слои. Группа включает несколько видов тканей, различающихся морфологическими и функциональными признаками.
4. Нервная (нейральная) ткань характеризуется способностью к возбудимости и проведению нервного импульса. Она образована (а) собственно нервными клетками (нейронами) отростчатой формы, связанными друг с другом в цепи и сложные системы посредством специализированных соединений (синапсов), и (б) клетками, осуществляющими вспомогательные функции - нейроглией. Основная функция нервной ткани - интеграция отдельных частей организма и регуляция его функций. Критерии объединения тканей в каждую из четырех указанных выше групп не полностью идентичны: при выделении эпителиальных и соединительных тканей за основу принимались преимущественно морфологические признаки, при определении специфики мышечных и нервной тканей исходили, главным образом, из функциональных критериев. Каждая группа (кроме последней) включает ряд тканей, различающихся источниками своего эмбрионального развития. Гистогенетическая классификация тканей (наиболее известные ее варианты разработаны Н.Г. Хлопиным и В.П. Михайловым) основывается на происхождении тканей в процессах онто- и филогенеза. Она вскрывает глубинные гистогенетические связи между морфологически и функционально различными тканями, происходящими из одного эмбрионального зачатка. Эти связи и общие признаки, не всегда заметные в физиологических условиях жизнедеятельности тканей, могут ярко проявляться в процессах их регенерации, реактивных изменений или злокачественного роста. Универсальная классификация, охватывающая все тканевые типы, нуждается в уточнении и находит использование преимущественно у специалистов. Более широкое распространение получили гистогенетические классификации отдельных групп тканей (в частности, эпителия, мышечных тканей). Поскольку морфофункциональная и гистогенетическая классификации тканей дополняют друг друга, наиболее полная оценка свойств тканей должна учитывать как их монофункциональные, так и гистогенетические характеристики.
Тема 2. ЭПИТЕЛИАЛЬНЫЕ ТКАНИ ОБЩИЕ СВЕДЕНИЯ Эпителиальные ткани, или эпителии (от греч. epi - над и thele - сосок, тонкая кожица) - пограничные ткани, которые располагаются на границе с внешней средой, покрывают поверхность тела, выстилают его полости, слизистые оболочки внутренних органов и образуют большинство желез. Различают три вида эпителиев: 1) покровные эпителии (образуют разнообразные выстилки), 2) железистые эпителии (образуют железы), 3) сенсорные эпителии (выполняют рецепторные функции, входят в состав органов чувств). Функции эпителиев: 1. Разграничительная, барьерная - основная функция эпителиев, все остальные являются ее частными проявлениями. Эпителии образуют барьеры между внутренней средой организма и внешней средой; свойства этих барьеров (механическая прочность, толщина, проницаемость и др.) определяются конкретными структурно-функциональными особенностями каждого эпителия. Немногими исключениями из общего правила служат эпителии, разграничивающие две области внутренней среды - например, выстилающие полости тела (мезотелий) или сосуды (эндотелий). 2. Защитная - эпителии обеспечивают защиту внутренней среды организма от повреждающего действия механических, физических (температурных, лучевых), химических и микробных факторов. Защитная функция может выражаться по-разному (например, эпителии могут образовывать толстые пласты, формировать наружный малопроницаемый, физически и химически устойчивый роговой слой, секретировать защитный слой слизи, вырабатывать вещества, обладающие антимикробным действием, и др.). 3. Транспортная - может проявляться переносом веществ сквозь пласты эпителиальных клеток (например, из крови через эндотелий мелких сосудов в окружающие ткани) или по их поверхности (например, транспорт слизи мерцательным эпителием дыхательных путей или ооцита мерцательным эпителием маточной трубы). Вещества могут переноситься через эпителиальный пласт механизмами диффузии, транспорта, опосредованного белками-переносчиками, и везикулярного транспорта. 4. Всасывающая - многие эпителии активно всасывают вещества; наиболее яркими их примерами служат эпителии кишки и почечных канальцев. Эта функция, по сути, представляет собой частный вариант транспортной функции. 5. Секреторная - эпителии являются функционально ведущими тканями большей части желез. 6. Экскреторная - эпителии участвуют в удалении из организма (с мочой, потом, желчью и др.) конечных продуктов обмена веществ или введенных в организм (экзогенных) соединений (например, лекарств). 7. Сенсорная (рецепторная) - эпителии, находясь на границе внутренней среды организма и внешней среды, воспринимают сигналы (механические, химические), исходящие из последней. СТРОЕНИЕ РАЗЛИЧНЫХ ВИДОВ ЭПИТЕЛИЯ Однослойные эпителии - эпителиальные ткани, все клетки которых располагаются на базальной мембране. По форме образующих их клеток они подразделяются на плоские, кубические или призматические. Призматические эпителии могут быть однорядными, если ядра их клеток располагаются на одном уровне, и многорядными (псевдомногослойными), если (вследствие различной формы и высоты клеток) ядра лежат на разных уровнях.
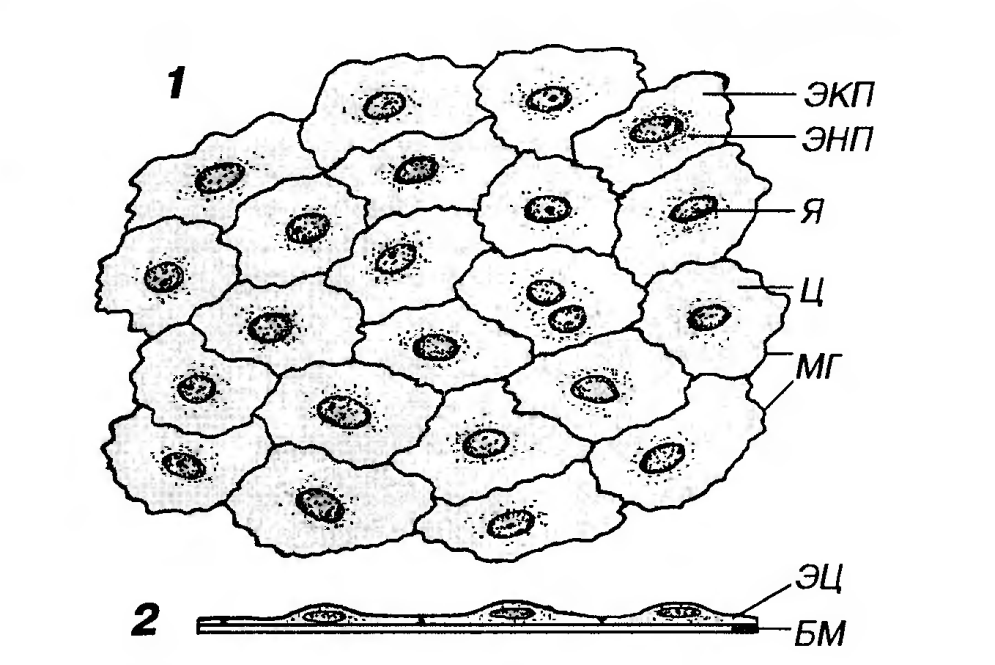
1. Однослойный плоский эпителий образован уплощенными клетками с некоторым утолщением в области расположения дисковид-ного ядра (рис. 4-5). Этим клетками свойственна диплазматическая дифференцировка цитоплазмы: она подразделяется на внутреннюю часть (эндоплазму), которая располагается вокруг ядра и содержит большую часть сравнительно немногочисленных органелл, и наружную часть (эктоплазму), относительно свободную от органелл. Вследствие малой толщины эпителиального пласта через него легко диффундируют газы и быстро транспортируются различные метаболиты. Примерами такого эпителия служат выстилки сосудов - эндотелий, полостей тела - мезо-телий (входит в состав серозных оболочек), некоторых почечных канальцев (тонкая часть петли Генле), альвеол легкого (клетки I типа). Камбиальные элементы в таком эпителии располагаются диффузно.
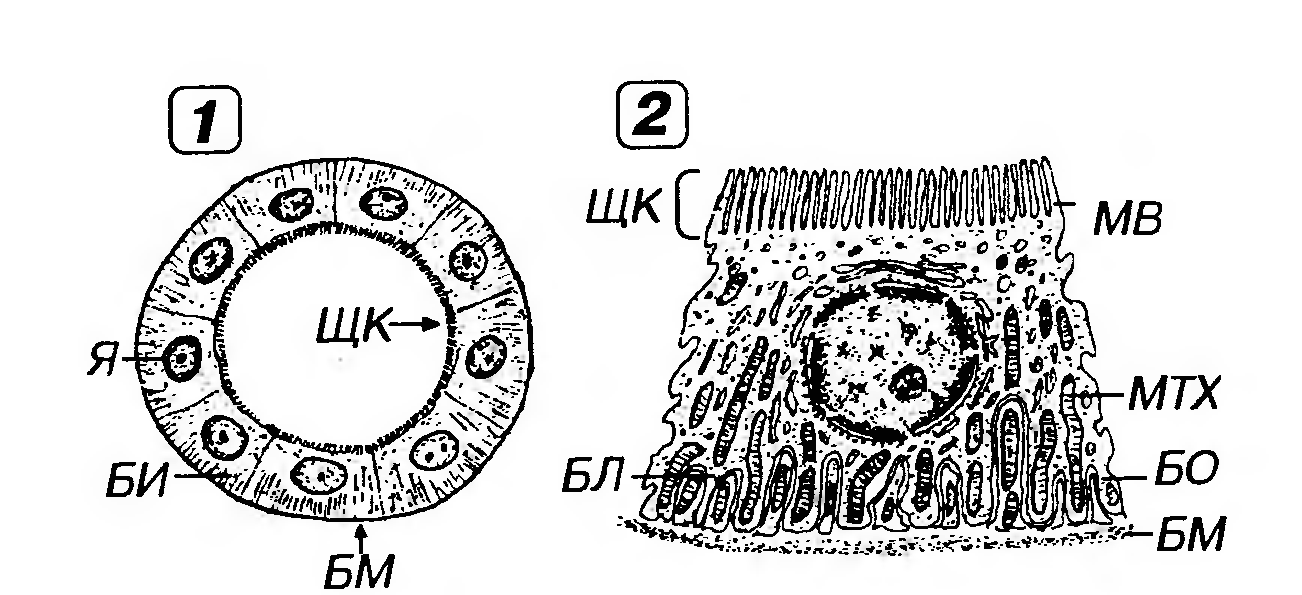
Рис. 5. Однослойный плоский эпителий (мезотелий сальника). 1 - вид с поверхности (пленочный препарат); 2 - вид на поперечном срезе. Я - ядро; Ц - цитоплазма; ЭНП - эндоплазма; ЭКП - эктоплазма, МГ - межклеточные границы (выявлены импрегнацией нитратом серебра); ЭЦ - эпителиоциты; БМ - базальная мембрана. 2. Однослойный кубический эпителий образован клетками, содержащими ядро сферической формы и набор органелл, которые развиты лучше, чем в клетках плоского эпителия. Такой эпителий встречается в почечных канальцах (рис. 6, см. также рис. 2), в которых он имеет базальную исчерченность, а в части канальцев - и щеточную каемку (однослойный кубический каемчатый эпителий, участвующий в процессах всасывания), в фолликулах щитовидной железы, в мелких протоках поджелудочной железы, желчных протоках печени, мелких собирательных трубочках почки. Камбий этого эпителияобычно диффузный (за исключением образующего протоки).
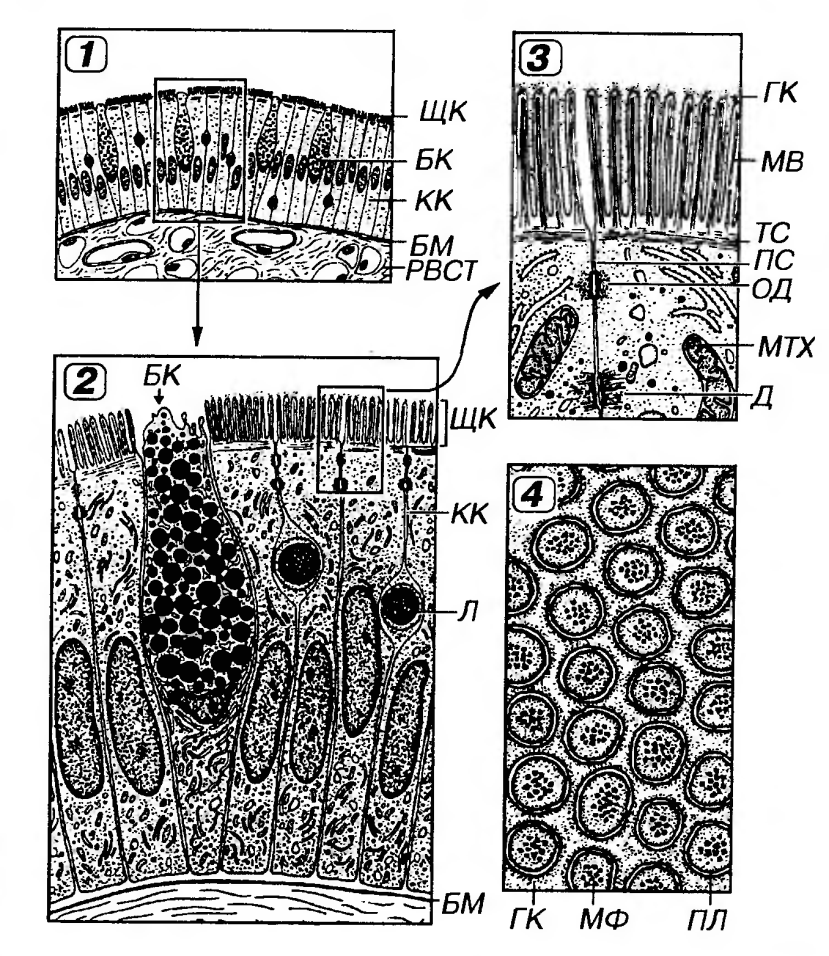
Рис. 6. Однослойный кубический каемчатый эпителий (почечный каналец, проксимальный отдел нефрона). 1 - поперечное сечение канальца, 2 - эпителиоцит. Я - ядро, БИ - базальная исчерченность, БМ - базальная мембрана, ЩК - щеточная каемка, MB - микроворсинки, БЛ - базальный лабиринт, БО - базальные отростки, MTX - митохондрии. 3. Однослойный призматический (цилиндрический, или столбчатый) эпителий образован клетками с резко выраженной полярностью. Ядро эллипсоидной формы лежит вдоль длинной оси клеток и обычно несколько смещено к их базальной части, а хорошо развитые органеллы неравномерно распределены по цитоплазме. Такой эпителий покрывает поверхность желудка, кишки, образует выстилку крупных протоков поджелудочной железы, крупных желчных протоков, желчного пузыря, маточной трубы, стенку крупных собирательных трубочек почки. В кишке и желчном пузыре этот эпителий каемчатый.Для большинства указанных эпителиев характерны функции секреции и (или) всасывания. Так, в эпителии тонкой кишки (рис. 7), встречаются два основных типа дифференцированных клеток - призматические каемчатые (обеспечивающие пристеночное пищеварение и всасывание) и бокаловидные (вырабатывающие слизь, которая выполняет защитную функцию). Камбий в указанных эпителиях, как правило, локализованный (например, в кишечных криптах или шейке желез желудка). По мере дифференцировки клетки приобретают характерные специфические признаки и смещаются из области локализации камбиальных элементов в зону расположения зрелых клеток. В таком однослойном эпителиальном пласте клетки различных участков обладают неодинаковым строением и функциями, что обозначается как горизонтальная анизоморфия.
Рис. 7. Однослойный призматический каемчатый эпителий (тонкая кишка) 1 - общий вид под световым микроскопом, 2 - ультраструктурная организация 3 - апикальная часть эпителиоцита, 4 - поперечный разрез микроворсинок КК - каемчатая клетка, ЩК - щеточная каемка, БК - бокаловидная клетка, Л - лимфоцит (внутризпителиальный), БМ - базальная мембрана. РВСТ - рыхлая волокнистая соединительная ткань, MB - микроворсинки, ГК - гликокаликс. МФ - микрофиламенты ПЛ - плазмолемма, ТС - терминальная сеть (актиновых микрофиламентов)' ПС – плотное соединение, ОД - опоясывающая десмосома, Д - десмосома, MTX - митохондрия. Однослойный многорядный (псевдомногослойный) призматический эпителий образован клетками нескольких типов, имеющих различные размеры. В этих клетках ядра располагаются на разных уровнях, что создает ложное впечатление многослойности (обусловливая второе название эпителия).
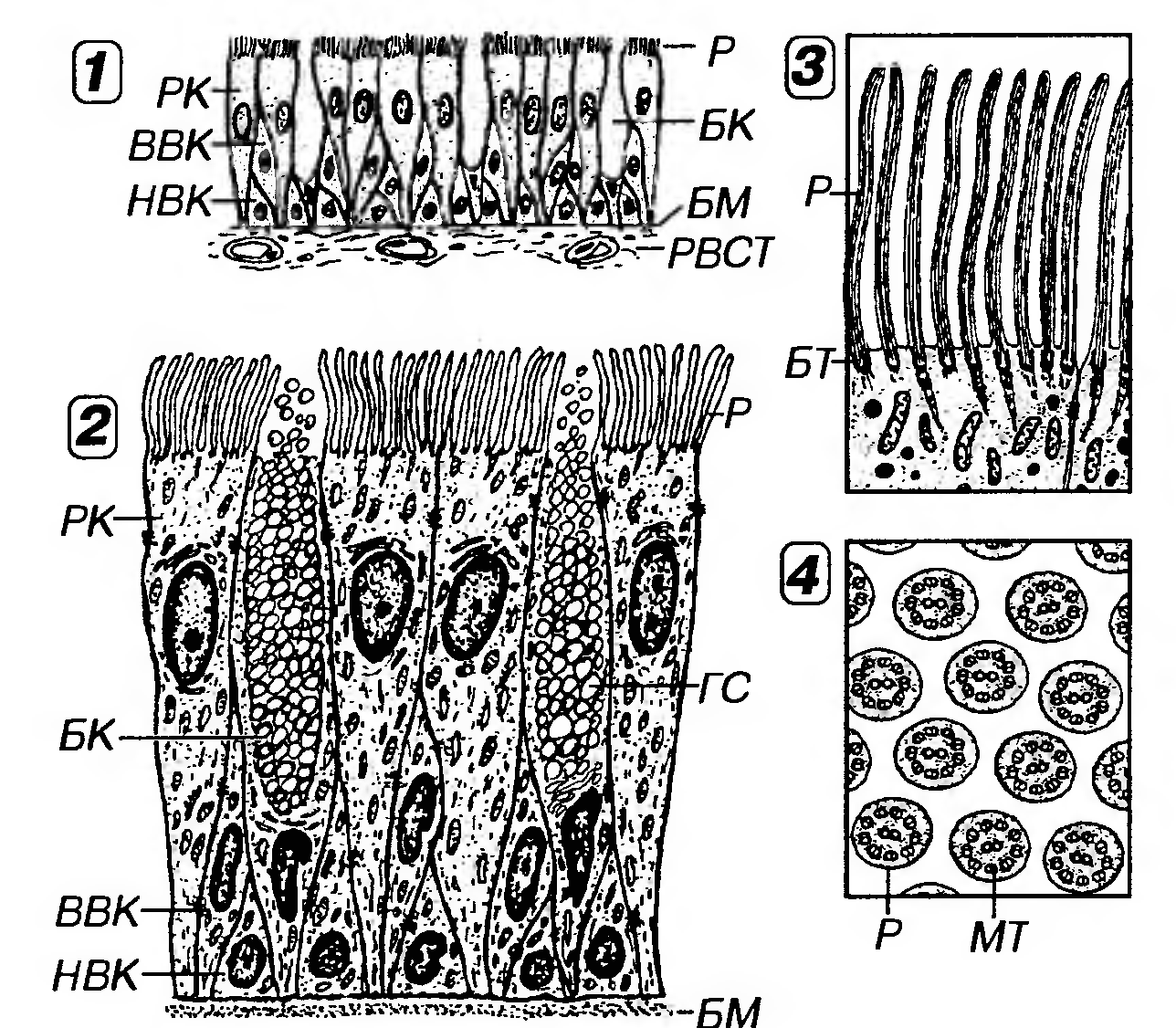
Рис. 8. Однослойный многорядный призматический реснитчатый (мерцательный) эпителий воздухоносных путей. 1 - общий вид под световым микроскопом. 2 - ультраструктурная организация, 3 - апикальная часть эпителиоцита, 4 - поперечный разрез ресничек. РК - реснитчатые клетки, Р - реснички, БК - бокаловидная клетка. ГС - глобулы слизи, ВВК - высокая вставочная клетка, НВК - низкая вставочная клетка, БМ - базальная мембрана, PBCT - рыхлая волокнистая соединительная ткань, МТ - микротрубочки, БТ - базальное тельце. Однослойный многорядный призматический реснитчатый (мерцательный) эпителий воздухоносных путей - наиболее типичный представитель многорядных эпителиев (рис. 8). В нем имеются клетки четырех основных типов: (1) низкие вставочные (базальные), (2) высокие вставочные (промежуточные), (3) реснитчатые (мерцательные) и (4) бокаловидные. Камбиальными элементами служат низкие вставочные клетки, которые имеют мелкие размеры. Своим широким основанием они прилежат к базальной мембране, обусловливая прикрепление к ней всего эпителиального пласта, а узкой апикальной частью не доходят до просвета. Наиболее дифференцированные клетки эпителия - реснитчатые (численно преобладают) и бокаловидные. Последние вырабатывают слизь, которая покрывает поверхность эпителия, перемещаясь по ней благодаря биению ресничек мерцательных клеток. Реснитчатые и бокаловидные клетки своей узкой базальной частью контактируют с базальной мембраной и прикрепляются к вставочным клеткам, а апикальной - граничат с просветом органа.
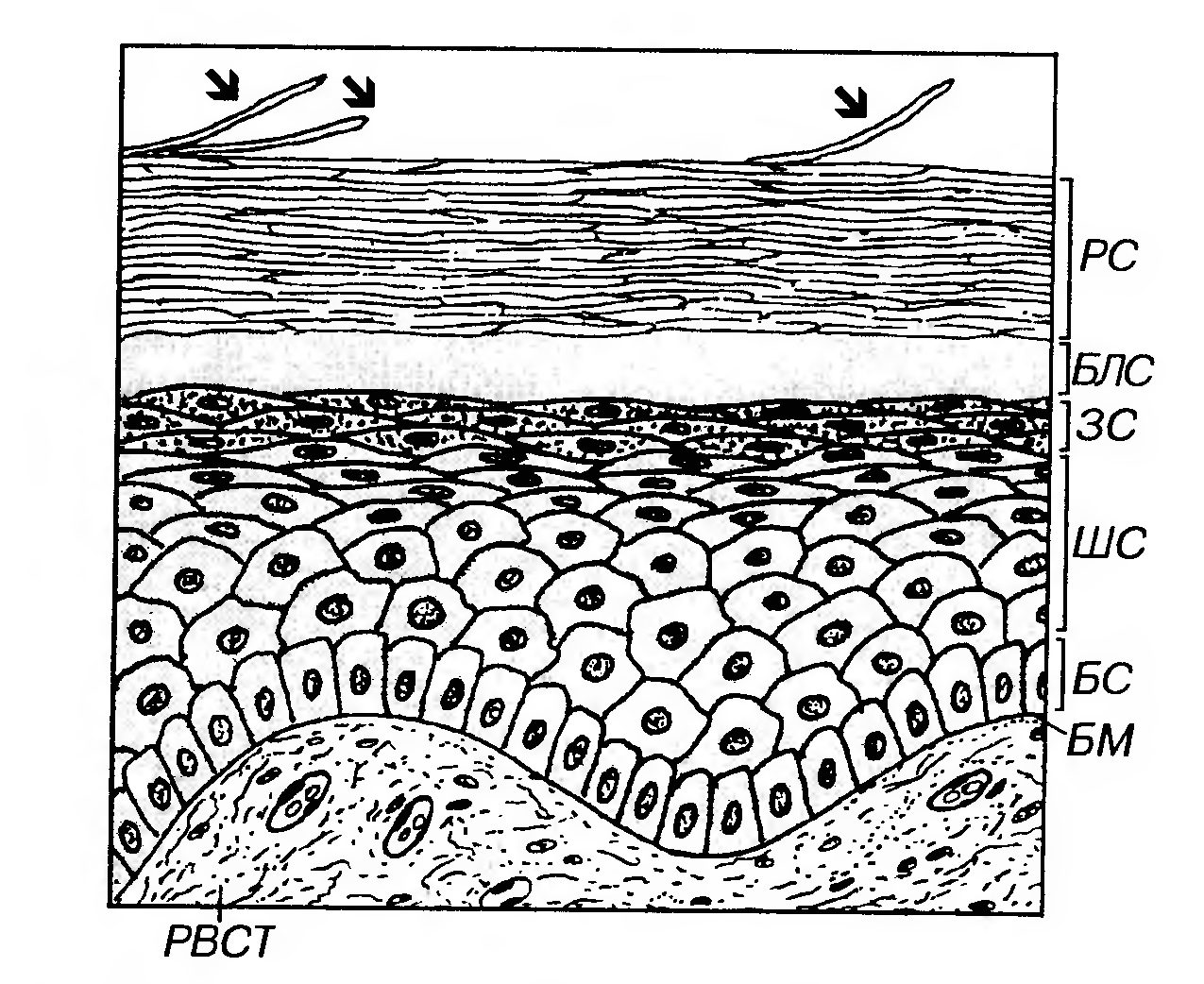
Однослойный двурядный призматический эпителий встречается в протоке придатка яичка, семявыносящем протоке, концевых отделах предстательной железы, семенных пузырьков. Многослойные эпителии - эпителии, в которых лишь часть клеток (образующих базальный слой) располагается на базальной мембране; клетки, входящие в состав остальных слоев, утрачивают с ней связь. Форма клеток в различных слоях таких эпителиев неодинакова; в целом форму всего эпителиального пласта оценивают по форме клеток поверхностного слоя. Морфологическая классификация многослойных эпителиев выделяет: (1) многослойные плоские эпителии, (2) многослойные кубические эпителии, (3) многослойные призматические эпителии и (4) переходный эпителий - особый вид многослойного эпителия, форма клеток которого непостоянна и изменяется в зависимости от функционального состояния органов, которые он выстилает (см. ниже). Многослойные плоские эпителии - наиболее распространенный вид многослойных эпителиев в организме человека. Они, в свою очередь, подразделяются (в зависимости от наличия или отсутствия рогового слоя) на (а) ороговевающие и (б) неороговевающие. Поддержание целостности многослойных эпителиев обеспечивается тем, что эпителиоциты непрерывно образуются в самом глубоком (базальном) слое благодаря делению малодифференцированных камбиальных клеток, затем смешаются в вышележащие слои, подвергаются дифференцировке и в конечном итоге слущиваются (десквамируют) с поверхности пласта. Процессы пролиферации и дифференцировки эпителиальных клеток регулируются рядом биологически активных веществ, часть которых выделяется клетками подлежащей соединительной ткани. Наиболее важными из них являются цитокины, в частности, эпидермальный фактор роста (ЭФР), интерлейкины (ИЛ) -1 и -6, инсулиноподобные факторы роста I и II, трансформирующий фактор роста-α (ТФРα), а также витамины А и D. В гормонально-зависимых органах на них влияют соответствующие гормоны. Дифференцировка эпителиоцитов сопровождается изменением экспрессии синтезируемых ими цитокератинов, образующих промежуточные филаменты. Клиническое значение изучения факторов, регулирующих пролиферацию и дифференцировку эпителиоцитов, связано с выявлением их роли в патогенезе различных заболеваний, а также с возможностью их применения для воздействия на ткани in vivo и in vitro. Так, благодаря методам тканевой инженерии путем стимуляции развития фрагментов кожного эпителия в условиях in vitro в течение сравнительно короткого времени удается получить его пласты, достаточные по площади для аутотрансплантации (пересадке в пределах собственного организма), например, при обширных ожогах, язвах и пролежнях. Десквамация клеток (в ороговевающем эпителии - роговых чешуек) с поверхности эпителиального пласта выполняет роль важного защитного механизма многослойных эпителиев, так как она обеспечивает постоянное удаление прикрепившихся к ним патогенных микроорганизмов, потенциально способных осуществить внедрение (инвазию) в эпителий и подлежащие ткани. Механизмом десквамации удаляются и заменяются новыми наиболее поверхностные клетки, поврежденные механически и химически и уже неспособные выполнять барьерную функцию. Многослойный плоский ороговевающий эпителий образует наружный слой кожи (эпидермис), покрывает поверхность некоторых участков слизистой оболочки полостирта. Он состоит из пяти слоев: (1) базального, (2) шиповатого, (3) зернистого (4) блестящего и (5) рогового (рис. 9). 1) Базальный слой образован клетками кубической или призматической формы, лежащими на базальной мембране. Для них характерно овальное ядро с одним или двумя ядрышками и базофильная цитоплазма, содержащая хорошо развитые органеллы, многочисленные промежуточные кератшовые филаменты (тонофиламенты). Основные функции базального слоя: а) содержит камбиальные элементы эпителия; б) обеспечивает прикрепление эпителия к подлежащей соединительной ткани (его клетки связаны с соседними эпителиоцитами десмосомами, а с базальной мембраной - полудесмосомами). Помимо десмосом между эпителиоцитами имеются щелевые и плотные соединения.
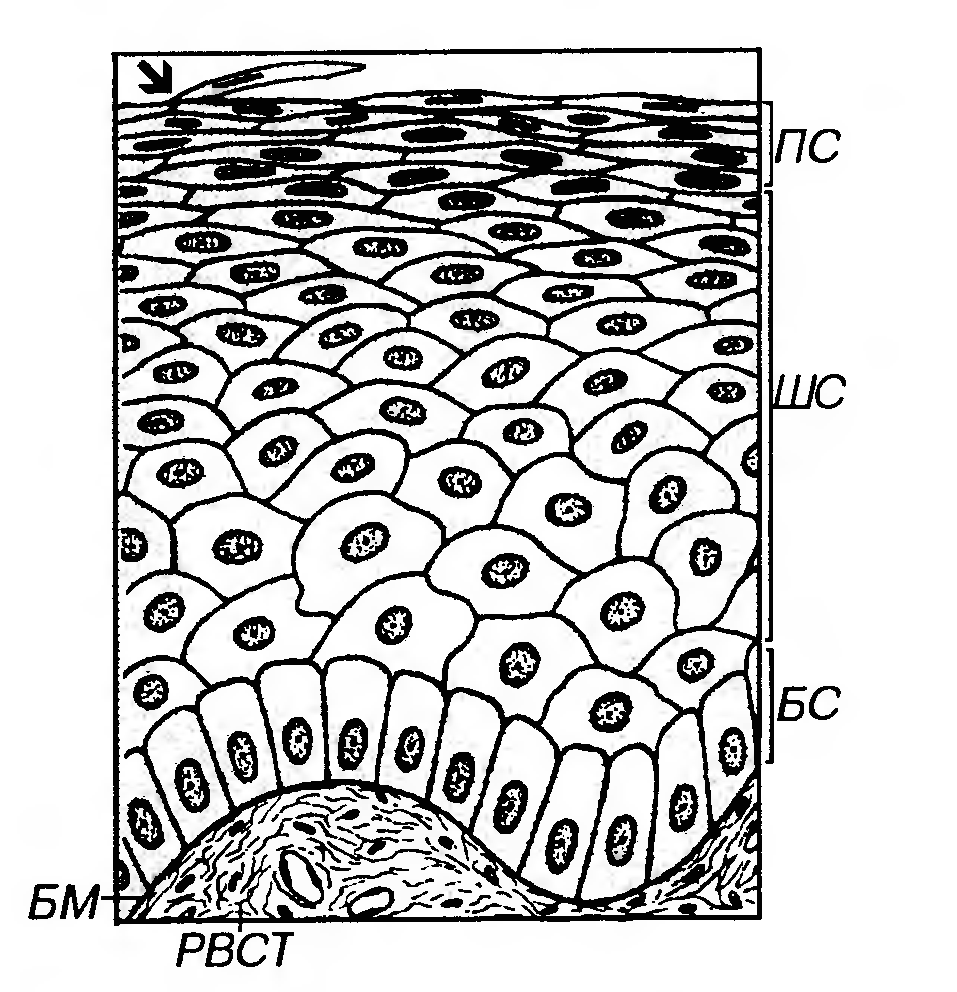
Рис. 9. Многослойный плоский ороговевающий эпителий (эпидермис) - вид на гистологическом препарате. БМ - базальная мембрана, БС - базальный слой, ШС - шиповатый слой, ЗС - зернистый слой, БЛС - блестящий слой, PC - роговой слой. Десквамирующие роговые чешуйки показаны стрелками. 2) Шиповатый слой образован крупньши клетками неправильной формы, связанными друг с другом десмосомами в области многочисленных отростков ("шипов"), которые содержат пучки тонофиламентов. В глубоких частях слоя могут встречаться отдельные делящиеся клетки. По мере приближения к зернистому слою клетки из полигональных постепенно становятся уплощенными. 3) Зернистый слой - сравнительно тонкий, образован уплощенными (веретеновидньши на разрезе) клетками. Ядро - плоское, с конденсированным хроматином, цитоплазма содержит многочисленные тонофиламенты и гранулы двух типов: а) кератогиалиновые - крупные (0.5-1 мкм), базофильные (электронно-плотные), неправильной формы, содержащие профилаггрин - важный компонент, необходимый для последующего образования рогового вещества (кератина). Профилаггрин является предшественником филаггрина - белка, организующего агрегацию кератиновых промежуточных филаментов в крупные комплексы - макрофибриллы. Он служит своеобразным матриксом, в который погружаются тонофиламенты. б) пластинчатые (кератиносомы) - мелкие, удлиненные, размером около 250 нм (видны только под электронным микроскопом), с пластинчатой структурой. Образуются еще в наружных отделах шиповатого слоя. Содержат ряд ферментов и липидов, которые при экзоцитозе гранул (в наружных отделах зернистого слоя) выделяются в межклеточное пространство, обеспечивая барьерную функцию и водонепроницаемость эпителия. По мере приближения к роговому слою клетки зернистого слоя претерпевают резкие изменения, подвергаясь ороговению (см. ниже). 4) Блестящий слой выражен только в эпителии толстой кожи, покрывающей ладони и подошвы. Он представляет собой зону перехода от живых клеток зернистого слоя к чешуйкам рогового слоя, не обладающим признаками живых клеток. На гистологических препаратах он имеет вид узкой оксифильной гомогенной полоски и состоит из уплощенных клеток, превращающихся в роговые чешуйки. На электронно-микроскопическом уровне этот слой обычно не выделяется. В блестящем слое завершаются процессы ороговения (в эпителии тонкой кожи, где этот слой не выражен, они происходят при переходе из зернистого слоя в роговой). Процесс ороговения эпителия заключается в превращении его живых эпителиальных клеток в роговые чешуйки - механически прочные и химически устойчивые постклеточные структуры, образующие в совокупности роговой слой эпителия, который обладает защитными свойствами. Хотя собственно формирование роговых чешуек происходит в наружных отделах зернистого слоя или в блестящем слое, синтез веществ, обеспечивающих ороговение, осуществляется еще в шиповатом слое. Основные процессы, происходящие в клетках в ходе ороговения: (1) изменение формы - клетки резко уплощаются, приобретая шестиугольную форму; (2) сборка и стабилизация пучков кератиновых промежуточных филаментов, образующих сети в цитоплазме и составляющих 80% ее массы. Процесс организации сетей филаментов с формированием их крупных пучков (макрофиламентов) обеспечивается филаггрином, который по его завершении разрушается. Последующая стабилизация системы филаментов осуществляется путем формирования межмолекулярных дисульфидных связей. Кератиновые филаменты присутствуют в клетках, начиная с базального слоя, филаггрин в виде неактивного предшественника (профилаггрина) впервые обнаруживается в кератогиалиновых гранулах зернистого слоя; (3) образование оболочки роговой чешуйки - плотной, механически прочной и химически устойчивой белковой структуры - происходит путем упорядоченного отложения ряда белков (инволюкрина, цистатина-α, лорикрина и др.) на внутренней поверхности плазмолеммы с образованием между ними многочисленных ковалентных связей. Сборка оболочки роговой чешуйки катализируется ферментом трансглутаминазой (который, как и инволюкрин - его главный субстрат и основной компонент оболочки - впервые обнаруживается в шиповатом слое). (4) полное ферментное разрушение остальных структур цитоплазмы и ядра; (5) дегидратация цитоплазмы (с потерей 70% массы клетки). 5) Роговой слой - наиболее поверхностный - имеет максимальную толщину в эпителии кожи (эпидермисе) в области ладоней и подошв. Он образован плоскими роговыми чешуйками с резко утолщенной плазмолеммой (оболочкой), не содержащими ядра и органелл и заполненными сетью из толстых пучков кератиновых филаментов, погруженных в плотный матрикс. Роговые чешуйки в течение определенного времени сохраняют связи друг с другом и удерживаются в составе пластов благодаря частично сохраненным десмосомам, а также взаимному проникновению бороздок и гребешков, образующих ряды на поверхности соседних чешуек. В наружных частях слоя десмосомы полностью разрушаются, и роговые чешуйки слущиваются с поверхности эпителия. Многослойный плоский неороговевающий эпителий покрывает поверхность роговицы глаза, конъюнктивы, слизистых оболочек полости рта (частично), глотки, пищевода, влагалища, влагалищной части шейки матки, части мочеиспускательного канала. Он образован тремя слоями клеток: (1) базальным, (2) шиповатым (промежуточным) и (3) поверхностным (рис. 10).
Рис. 10. Многослойный плоский неороговевающий эпителий (влагалище) - вид на гистологическом препарате. БМ - базальная мембрана, БС - базальный слой, ШС - шиповатый слой, ПС - поверхностный слой. Десквамирующая клетка поверхностного слоя показана стрелкой. 1) Базальный слой аналогичен по строению и функции соответствующему слою ороговевающего эпителия. 2) Шиповатый (промежуточный) слой образован крупными полигональными клетками, которые по мере приближения к поверхностному слою уплощаются. Их цитоплазма заполняется многочисленными тонофиламентами, которые располагаются в ней диффузно, не образуя крупных пучков. В клетках наружных отделов этого слоя накапливается керотогиалин в виде мелких округлых гранул. 3) Поверхностный слой нерезко отделен от шиповатого. Он образован уплощенными клетками, содержащими рыхло распределенные цитокератиновые филаменты, которые по химическому составу отличаются от таковых в роговых чешуйках. Содержание органелл снижено по сравнению с таковым в клетках шиповатого слоя, плазмолемма утолщена, межклеточные пространства редуцированы. Ядро - светлое (везикулярное) или темное, с плохо различимыми гранулами хроматина (пикнотическое). Механизмом десквамации клетки этого слоя постоянно удаляются с поверхностности эпителия. Многослойный кубический эпителий в организме человека встречается редко. Он сходен по строению с многослойным плоским эпителием, но клетки поверхностного слоя в нем имеют кубическую форму. Такой эпителий образует стенку крупных фолликулов яичника, выстилает протоки потовых и сальных желез кожи. Многослойный призматический эпителий, как и многослойный кубический, у человека встречается редко. Он выстилает некоторые участки мочеиспускательного канала, крупные выводные протоки слюнных и молочных желез (частично). Такой эпителий обнаруживается в участках резкого перехода многослойного плоского эпителия в однослойный многорядный и образует узкую зону между этими эпителиями (например, в глотке и гортани). Переходный эпителий - особый вид многослойного эпителия, который выстилает большую часть мочевыводящих путей (чашечки, лоханки, мочеточники и мочевой пузырь, часть мочеиспускательного канала), отчего его называют также уротелием. Форма клеток этого эпителия и толщина всего эпителиального пласта зависят от функционального состояния (степени растяжения) органа. Переходный эпителий образован тремя слоями клеток: базальным, промежуточным и поверхностным (рис. 11).
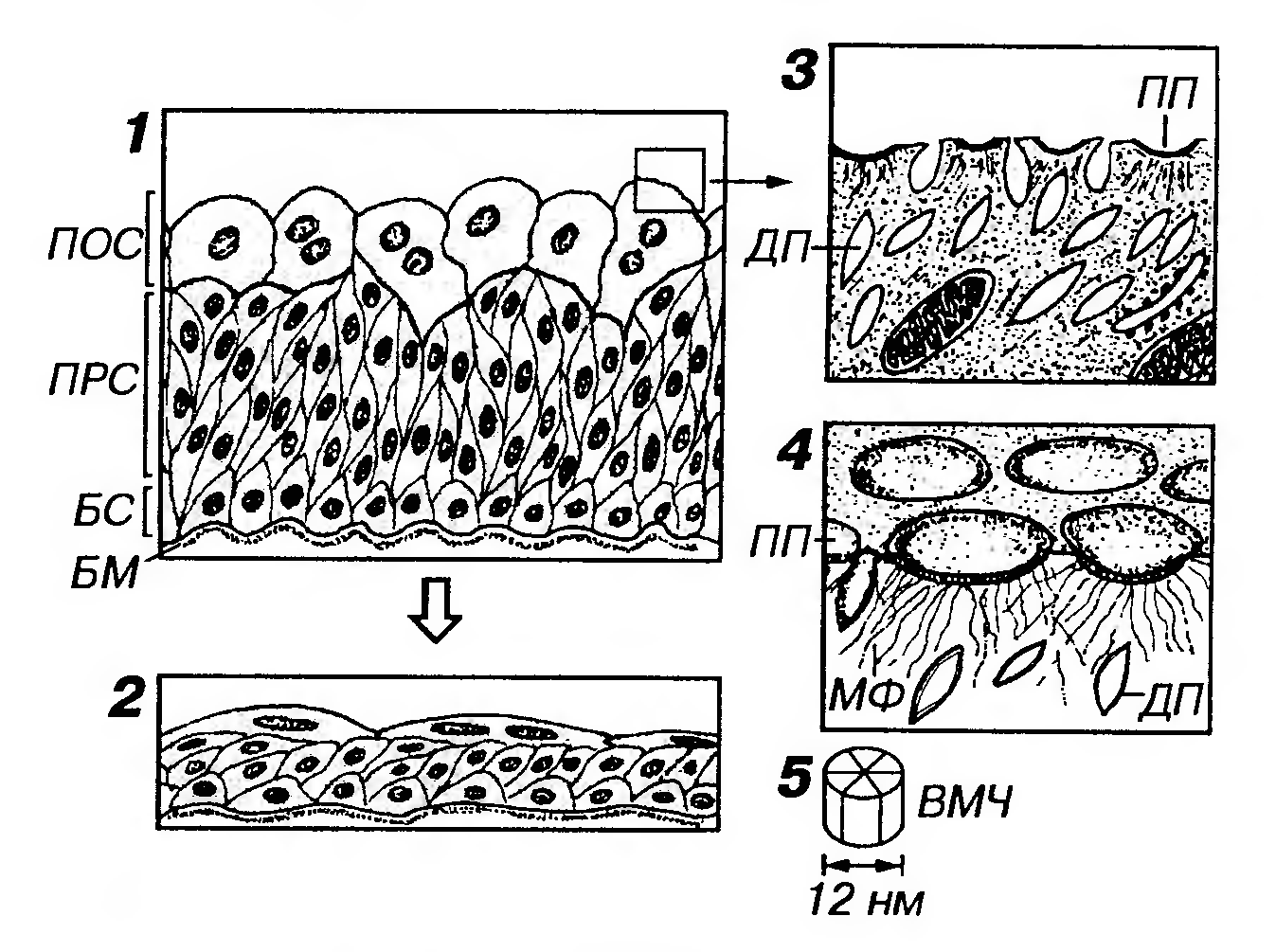
Рис. 11. Переходный эпителий. 1 - в состоянии покоя, 2 - в состоянии растяжения, 3 - участок цитоплазмы апикальной части фасеточной клетки на срезе, 4 - то же в объемном изображении, 5 - внутримембранная частица (ВМЧ), входящая в состав пластинок плазмолеммы (ПП) фасеточной клетки. БМ - базальная мембрана, БС - базальный слой, ПРС - промежуточный слой, ПОС - поверхностный слой (образован фасеточными клетками), ДП - дисковидные пузырьки, МФ - микрофиламенты. (1) базальный слой образован мелкими клетками, имеющими на срезе преимущественно треугольную форму и своим широким основанием прилежащими к тонкой базальной мембране; (2) промежуточный слой состоит из удлиненных клеток, более узкой частью направленных к базальному слою и черепицеобразно накладывающихся друг на друга; (3) поверхностный слой образован крупными одноядерными полиплоидными или двуядерными поверхностными (фасеточными) клетками, которые в наибольшей степени изменяют свою форму при растяжении эпителия (от округлой до плоской). Этому способствует формирование в апикальной части цитоплазмы этих клеток в состоянии покоя многочисленных инвагинаций плазмолеммы и особых дисковидных (ве-ретеновидных на срезах) пузырьков (длиной 0.3-0.8 мкм и шириной 0.12-0.18 мкм) - резервов плазмолеммы, которые встраиваются в нее по мере растяжения клетки. Формированию инвагинаций плазмолеммы способствуют многочисленные микрофиламенты, которые прикрепляются к ее особым участкам - пластинкам плазмолеммы. Пластинки плазмолеммы - утолщенные, сравнительно ригидные и малопроницаемые для воды полигональные участки апикальной плазмолеммы площадью 0.05-0.25 мкм2, которые в совокупности занимают до 75% ее поверхности. Пластинки содержат скопления внутримембранных белковых частиц размером около 12 нм, каждая из которых образована шестью субъединицами диаметром 5 нм. Пластинки плазмолеммы располагаются в виде "булыжной мостовой" (за тем лишь исключением, что каждая из них представляет собой не выпячивание, а вдавление на ее поверхности). Они разделены более гибкими участками плазмолеммы, не содержащими белковых частиц, которые способствуют образованию складок плазмолеммы. Мембрана, образующая пластинки плазмолеммы, собирается в комплексе Гольджи и транспортируется в апикальную плазмолемму посредством дисковидных пузырьков. Предполагают, что наличие указанных пластинок в апикальной плазмолемме поверхностных клеток и плотных соединений между латеральными поверхностями этих клеток обеспечивает непроницаемость переходного эпителия для воды. Это свойство данного эпителия имеет важнейшее функциональное значение, поскольку благодаря ему гипертоническая моча (накапливающаяся в мочевом пузыре) не разводится изотонической жидкостью из кровеносных сосудов подлежащей соединительной ткани собственной пластинки слизистой оболочки. Альтернативные представления о строении переходного эпителия основаны на данных некоторых исследователей, согласно которым клетки его промежуточного и даже поверхностного слоев своими тонкими отростками контактируют с базальной мембраной. В соответствии с такими представлениями, переходный эпителий следует считать не многослойным, а особым видом однойслойного многорядного. ЖЕЛЕЗЫ Железы выполняют секреторную функцию, вырабатывая и выделяя разнообразные продукты (секреты), обеспечивающие различные функции организма. Большинство желез образовано эпителиальной тканью (железистым эпителием), хотя той или иной способностью к секреции обладают все ткани. Классификация желез Существует несколько классификаций желез, которые основаны на учете различных признаков. Железы подразделяются: 1) по числу клеток (рис. 13) - на одноклеточные (например, бокаловидные клетки, клетки диффузной эндокринной системы) и многоклеточные (большинство желез); 2) по уровню организации - на входящие в состав различных органов в качестве их компонентов (например, железы слизистых оболочек) или являющиеся самостоятельными анатомически оформленными органами (например, крупные слюнные железы, печень, поджелудочная железа, щитовидная железа и др.); 3) по расположению (относительно эпителиального пласта) - на эндоэпителиальные и экзоэпителиальные, т.е. лежащие в пределах эпителиального пласта или вне его, соответственно (см. рис. 13). Большинство желез относится к экзоэпителиальным;
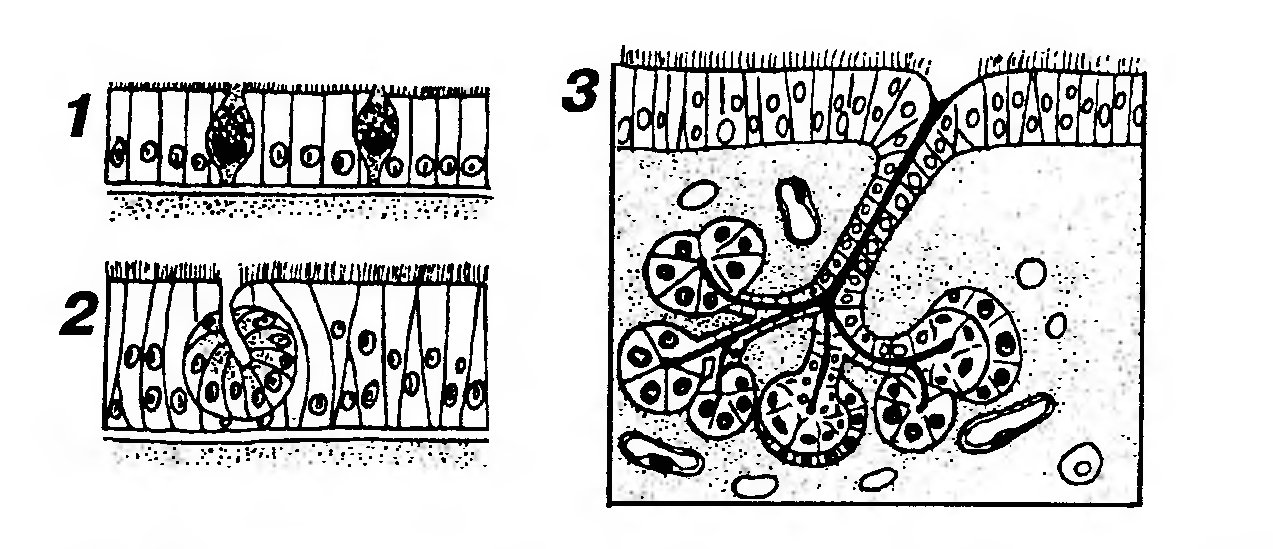
Рис. 13. Классификация экзокринных желез в зависимости от количества клеток, входящих в их состав, и расположения по отношению к эпителиальному пласту. 1 - одноклеточные эндоэпителиальные железы (бокаловидные клетки);2 - многоклеточная эндоэпителиальная железа (слизистой оболочки полости носа); 3 - многоклеточная экзоэпителиальная железа (слизистой оболочки трахеи или бронхов). 4) по месту (направлению) выведения секрета - на эндокринные (выделяющие секреторные продукты, называемые гормонами, в кровь) и экзокринные (выделяющие секреты на поверхность тела или в просвет внутренних органов); 5) по механизму (способу) выведения секрета (рис. 14) - на мерокринные (без нарушения структуры клетки), апокринные (с отделением в секрет части апикальной цитоплазмы) и голокринные (с полным разрушением клеток и выделением их фрагментов в секрет). В организме человека большинство желез относится к мерокринным; апокринных желез немного (например, часть потовых и молочные), к голокринным относятся лишь сальные железы. В клетках некоторых желез выведение секрета осуществляется одновременно двумя механизмами - апокринным и мерокринным. 6) по химическому составу вырабатываемого секрета – на белковые (серозные), слизистые, смешанные (белково-слизистые), липидные и др. Характеристики (5) и (6) используются преимущественно в отношении экзокринных желез.
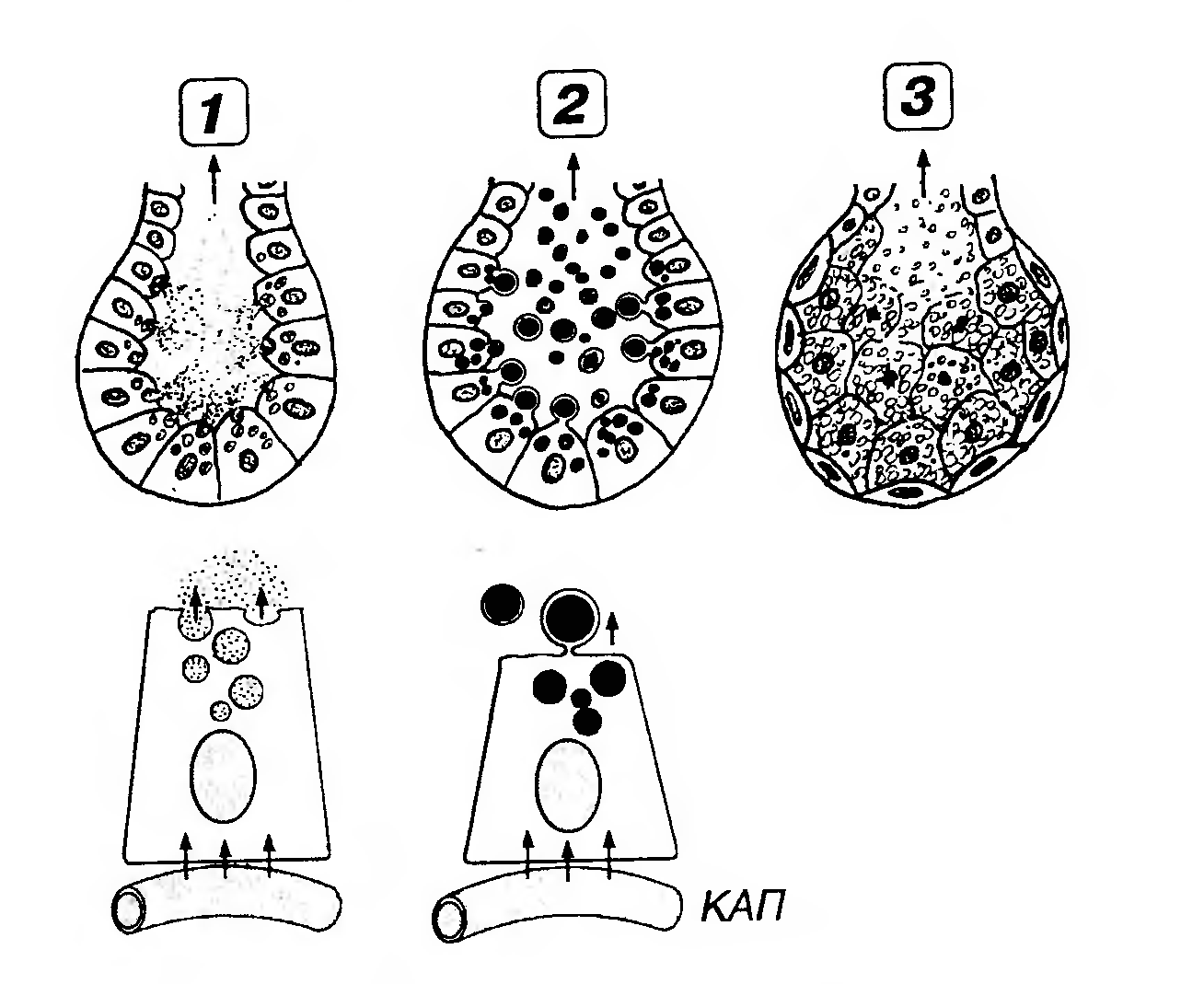
Рис. 14. Классификация экзокринных желез в зависимости от механизмов выведения секрета из гландулоцитов. 1 - мерокринная железа (секреция осуществляется без нарушения структуры клетки): секреторные продукты, накопившиеся в гранулах, выводятся из клеток после слияния мембраны гранул с плазмолеммой апикальной части клетки; 2 - апокринная железа (секреция осуществляется с отделением в секрет части апикальной цитоплазмы, содержащей накопленный секреторный продукт). Под рисунками концевых отделов показаны железистые клетки, захватывающие вещества, необходимые для синтеза секрета, из капилляров (КАП) и выделяющие готовые продукты механизмами мерокринной или апокринной секреции, соответственно; 3 - голокринная железа (секреция осуществляется с полным разрушением клеток и выделением их фрагментов в секрет). Убыль зрелых клеток вследствие цитолиза уравновешивается активным размножением камбиальных клеток, расположенных на базальной мембране. Эндокринные железы (железы внутренней секреции) продуцируют гормоны - вещества различной химической природы, циркулирующие в крови в н
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 241; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.111.125 (0.059 с.) |