
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биологическая роль углеводовСодержание книги
Поиск на нашем сайте
ХИМИЯ УГЛЕВОДОВ Впервые термин «углеводы» был предложен профессором Дерптского (ныне Тартуского) университета К.Г. Шмидтом в 1844 г. В то время предполагали, что все углеводы имеют общую формулу Cm(H2O)n, т.е. углерод + вода. Отсюда название «углеводы». Например, глюкоза и фруктоза имеют формулу С(Н2О)6, тростниковый сахар (сахароза) C12(H2O)11, крахмал [С6(Н2О)5]n и т.д. В дальнейшем оказалось, что ряд соединений, по своим свойствам относящихся к классу углеводов, содержат водород и кислород в несколько иной пропорции, чем указано в общей формуле (например, дезоксирибоза С5Н10О4). В 1927 г. Международная комиссия по реформе химической номенклатуры предложила термин «углеводы» заменить термином «глициды», однако старое название «углеводы» укоренилось и является общепризнанным. Химия углеводов занимает одно из ведущих мест в истории развития органической химии. Тростниковый сахар можно считать первым органическим соединением, выделенным в химически чистом виде. Произведенный в 1861 г. A.M. Бутлеровым синтез (вне организма) углеводов из формальдегида явился первым синтезом представителей одного из трех основных классов веществ (белки, липиды, углеводы), входящих в состав живых организмов. Химическая структура простейших углеводов была выяснена в конце XIX в. в результате фундаментальных исследований Э. Фишера. Значительный вклад в изучение углеводов внесли отечественные ученые А.А. Колли, П.П. Шорыгин, Н.К. Кочетков и др. В 20-е годы нынешнего столетия работами английского исследователя У. Хеуорса были заложены основы структурной химии полисахаридов. Со второй половины XX в. происходит стремительное развитие химии и биохимии углеводов, обусловленное их важным биологическим значением. БИОЛОГИЧЕСКАЯ РОЛЬ УГЛЕВОДОВ Углеводы наряду с белками и липидами являются важнейшими химическими соединениями, входящими в состав живых организмов. У человека и животных углеводы выполняют важные функции: энергетическую (главный вид клеточного топлива), структурную (обязательный компонент большинства внутриклеточных структур) и защитную (участие углеводных компонентов иммуноглобулинов в поддержании иммунитета). Углеводы (рибоза, дезоксирибоза) используются для синтеза нуклеиновых кислот, они являются составными компонентами нуклеотидных ко-ферментов, играющих исключительно важную роль в метаболизме живых существ. В последнее время все большее внимание к себе привлекают смешанные биополимеры, содержащие углеводы: гликопептиды и глико-протеины, гликолипиды и липополисахариды, гликолипопротеины и т.д. Эти вещества выполняют в организме сложные и важные функции. С нарушением обмена углеводов тесно связан ряд заболеваний: сахарный диабет, галактоземия, нарушение в системе депо гликогена, нетолерантность к молоку и т.д. Следует отметить, что в организме человека и животного углеводы присутствуют в меньшем количестве (не более 2% от сухой массы тела), чем белки и липиды; в растительных организмах за счет целлюлозы на долю углеводов приходится до 80% от сухой массы, поэтому в целом в биосфере углеводов больше, чем всех других органических соединений вместе взятых.
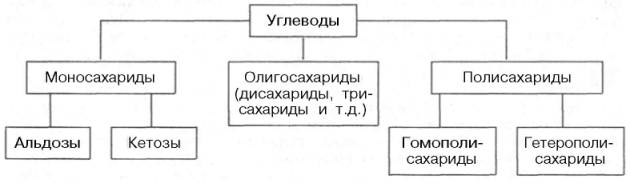
КЛАССИФИКАЦИЯ УГЛЕВОДОВ Углеводы можно определить как альдегидные или кетонные производные полиатомных (содержащих более одной ОН-группы) спиртов или как соединения, при гидролизе которых образуются эти производные. Согласно принятой в настоящее время классификации, углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды.
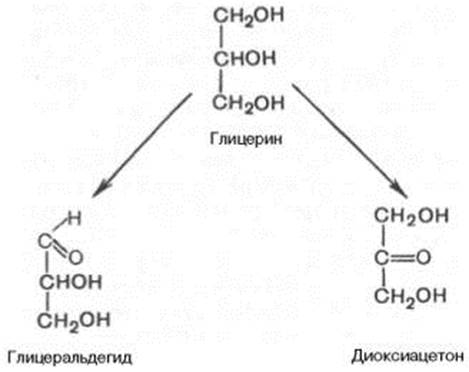
МОНОСАХАРИДЫ Моносахариды можно рассматривать как производные многоатомных спиртов, содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная группа находится в конце цепи, то моносахарид представляет собой альдегид и называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой. Простейшие представители моносахаридов – триозы: глицеральдегид и диоксиацетон. При окислении первичной спиртовой группы трехатомного спирта – глицерола – образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).
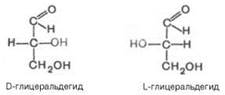
Стереоизомерия моносахаридов. Все моносахариды содержат асимметричные атомы углерода: альдотриозы – один центр асимметрии, альдо-тетрозы – 2, альдопентозы – 3, альдогексозы – 4 и т.д. Кетозы содержат на один асимметричный атом меньше, чем альдозы с тем же числом углеродных атомов. Следовательно, кетотриоза диоксиацетон не содержит асимметричных атомов углерода. Все остальные моносахариды могут существовать в виде различных стереоизомеров. Общее число стереоизомеров для любого моносахарида выражается формулой N = 2n, где N – число стереоизомеров, а n – число асимметричных атомов углерода. Как отмечалось, глицеральдегид содержит только один асимметричный атом углерода и поэтому может существовать в виде двух различных стереоизомеров. Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у асимметричного атома углерода расположена с правой стороны, принято считать D-глицеральдегидом, а зеркальное отражение – L-глицеральдегидом:
Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 16 стереоизомеров (24), представителем которых является, например, глюкоза. Для альдопентоз и альдотетроз число сте-реоизомеров равно соответственно 23 = 8 и 22 = 4. Все изомеры моносахаридов подразделяются на D- и L-формы (D-и L-конфигурация) по сходству расположения групп атомов у последнего центра асимметрии с расположением групп у D- и L-глицеральдегида. Природные гексозы: глюкоза, фруктоза, манноза и галактоза – принадлежат, как правило, по стереохимической конфигурации к соединениям D-ряда. Известно, что природные моносахариды обладают оптической активностью. Способность вращать плоскость поляризованного луча света – одна из важнейших особенностей веществ (в том числе моносахаридов), молекулы которых имеют асимметричный атом углерода или асимметричны в целом. Свойство вращать плоскость поляризованного луча вправо обозначают знаком плюс (+), а в противоположную сторону – знаком минус (–). Так, D-глицеральдегид вращает плоскость поляризованного луча вправо, т. е. D-глицеральдегид является D(+)-альдотриозой, а L-гли-церальдегид – L(–)-альдотриозой. Явление мутаротации имеет объяснение. Известно, что альдегиды и кетоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:
Реакция образования полуацеталя возможна и в пределах одной молекулы, если это не связано с пространственными ограничениями. По теории А. Байера, внутримолекулярное взаимодействие спиртовой и карбонильной групп наиболее благоприятно, если оно приводит к образованию пяти- или шестичленных циклов. При образовании полуацеталей возникает новый асимметрический центр (для D-глюкозы это С-1). Шестичленные кольца сахаров называют пиранозами, а пятичленные – фуранозами. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с α-модификацией моносахаридов D-ряда полуацетальный гидроксил пишут справа, а в формулах представителей L-ряда – слева. При написании β-формы поступают наоборот. Таким образом, явление мутаротации связано с тем, что каждый твердый препарат углеводов представляет собой какую-либо одну циклическую (полуацетальную) форму, но при растворении и стоянии растворов эта форма через альдегидную превращается в другие таутомерные циклические формы до достижения состояния равновесия. При этом значение удельного вращения, характерное для исходной циклической формы, постепенно меняется. Наконец, устанавливается постоянное удельное вращение, которое характерно для равновесной смеси таутомеров. Например, известно, что в водных растворах глюкоза находится главным образом в виде α- и β-глюкопираноз, в меньшей степени – в виде α- и β-глюкофураноз и совсем небольшое количество глюкозы – в виде альдегидной формы. Следует подчеркнуть, что из различных таутомерных форм глюкозы в свободном состоянии известны лишь α- и β-пиранозы. Существование малых количеств фураноз и альдегидной формы в растворах доказано, но в свободном состоянии они не могут быть выделены вследствие своей неустойчивости. Установлено, что величина удельного вращения при растворении любого моносахарида постепенно меняется и лишь при дли тельном стоянии раствора достигает вполне определенного значения. Например, для свежеприготовленного раствора глюкозы [α]D20= +112,2°, после длительного стояния раствора эта величина достигает равновесного значения [α]D20= +52,5°. Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией. Очевидно, мутаротация должна вызываться изменением асимметрии молекулы, а следовательно, трансформацией ее структуры в растворе. В 20-х годах У. Хеуорс предложил более совершенный способ написания структурных формул углеводов. Формулы Хеуорса – шести- или пятиугольники, причем они изображены в перспективе: кольцо лежит в горизонтальной плоскости. Находящиеся ближе к читателю связи изображают более жирными линиями (углеродные атомы цикла не пишут). Заместители, расположенные справа от остова молекулы при ее вертикальном изображении, помещают ниже плоскости кольца, а заместители, находящиеся слева,– выше плоскости кольца. Обратное правило применяют только для того единственного углеродного атома, гидроксильная группа которого участвует в образовании циклического полуацеталя. Так, у D-сахаров группу СН2ОН пишут над этим атомом углерода, а водородный атом при нем – внизу. Наконец, следует помнить, что при написании структурных формул по Хеуорсу гидроксильная группа при С-1 должна быть расположена ниже плоскости кольца в α-форме и выше – в β-форме:
Проекционные формулы Хеуорса не отражают подлинной конформации моносахаридов. Подобно циклогексану, пиранозное кольцо может принимать две конфигурации – форму кресла и форму лодки (конформацион-ные формулы). Форма кресла обычно более устойчива, и, по-видимому, именно она преобладает в большей части природных сахаров (рис. 5.1). ОЛИГОСАХАРИДЫ Олигосахариды – углеводы, молекулы которых содержат от 2 до 10 остатков моносахаридов, соединенных гликозидными связями. В соответствии с этим различают дисахариды, трисахариды и т.д. Дисахариды – сложные сахара, каждая молекула которых при гидролизе распадается на две молекулы моносахаридов. Дисахариды наряду с полисахаридами являются одними из основных источников углеводов в пище человека и животных. По строению дисахариды – это гликозиды, в которых 2 молекулы моносахаридов соединены гликозидной связью. Среди дисахаридов наиболее широко известны мальтоза, лактоза и сахароза. Мальтоза, являющаяся α-глюкопиранозил-(1–>4)-α-глюкопира-нозой, образуется как промежуточный продукт при действии амилаз на крахмал (или гликоген), содержит 2 остатка α-D-глюкозы (название сахара, полуацетальный гидроксил которого участвует в образовании гликозидной связи, оканчивается на «ил»).
В молекуле мальтозы у второго остатка глюкозы имеется свободный полуацетальный гидроксил. Такие дисахариды обладают восстанавливающими свойствами. Одним из наиболее распространенных дисахаридов является сахароза – обычный пищевой сахар. Молекула сахарозы состоит из одного остатка D-глюкозы и одного остатка D-фруктозы. Следовательно, это α-глюко-пиранозил-(1–>2)-β-фруктофуранозид:
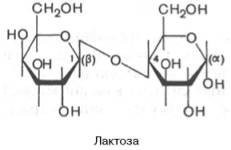
В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила и не обладает восстанавливающими свойствами. Гидролиз сахарозы приводит к образованию смеси, которую называют инвертированным сахаром. В этой смеси преобладает сильно левовращающая фруктоза, которая инвертирует (меняет на обратный) знак вращения правовращающего раствора исходной сахарозы. Дисахарид лактоза содержится только в молоке и состоит из D-галак-тозы и D-глюкозы. Это – β-галактопиранозил-(1–>4)-глюкопираноза:
Благодаря наличию в молекуле свободного полуацетального гидрокси-ла (в остатке глюкозы) лактоза относится к числу редуцирующих ди-сахаридов. Среди природных трисахаридов наиболее известна рафиноза, содержащая остатки фруктозы, глюкозы и галактозы. Рафиноза в больших количествах содержится в сахарной свекле и во многих других растениях. В целом олигосахариды, присутствующие в растительных тканях, разнообразнее по своему составу, чем олигосахариды животных тканей. ОЛИГО- И ПОЛИСАХАРИДЫ МЕТАБОЛИЗМ УГЛЕВОДОВ Гликолиз
Реакции гликолиза Сахара подвергаются метаболическим превращениям преимущественно в виде сложных эфиров фосфорной кислоты. Глюкоза, которую ткани получают из крови, в клетке также предварительно активируется путем фосфорилирования. В АТФ-зависимой реакции, катализируемой гексокиназой [1] глюкоза превращается в глюкозо-6-фосфат. После изомеризации глюкозо-6-фосфата в фруктозо-6-фосфат [2] последний вновь фосфорилируется с образованием фруктозо-1,6-дифосфата. Фосфофруктокиназа [3], катализирующая эту стадию, является важным ключевым ферментом гликолиза. До этого момента на одну молекулу глюкозы расходуются две молекулы АТФ. фруктозо-1,6-дифосфат расщепляется далее альдолазой [4] на два фосфорилированных С3-фрагмента. Эти фрагменты — глицеральдегид-3-фосфат и дигидроксиацетонфосфат — превращаются один в другой триозофосфатизомеразой [5]. Глицеральдегид-3-фосфат затем окисляется глицеральдегид-З-фосфатдегидрогеназой [6] с образованием НАДН + Н+. В этой реакции в молекулу включается неорганический фосфат с образованием 1,3-дифосфоглицерата. Такое промежуточное соединение содержит смешанную ангидридную связь, расщепление которой является высоко экзоэргическим процессом. На следующей стадии (катализируемой фосфоглицераткиназой [7]) гидролиз этого соединения сопряжен с образованием АТФ. Следующий промежуточный продукт, гидролиз которого может быть сопряжен с синтезом АТФ, образуется в реакции изомеризации 3-фосфоглицерата, полученного в результате реакции [7], в 2-фосфоглицерат (фермент: фосфоглицератмутаза [8]) и последующего отщепления воды (фермент: енолаза [9]). Продукт представляет собой сложный эфир фосфорной кислоты и енольной формы пирувата и потому называется фосфоенолпируватом (PEP). На последней стадии, которая катализируется пируваткиназой [10], образуются пируват и АТФ. Наряду со стадией [6] и тиокиназной реакцией в цитратном цикле это третья реакция, позволяющая клеткам синтезировать АТФ независимо от дыхательной цепи. Несмотря на образование АТФ она высоко экзоэргична и потому необратима. При гликолизе на активацию одной молекулы глюкозы потребляется 2 молекулы АТФ. В то же время при метаболическом превращении каждого С3-фрагмента образуются 2 молекулы АТФ. В результате выигрыш энергии составляет 2 моля АТФ на моль глюкозы. Изменение свободной энергии Энергетика метаболических процессов зависит не только от изменения стандартной свободной энергии ΔGo', но и от концентрации метаболита. На схеме представлены фактические изменения свободной энергии ΔG на отдельных стадиях гликолиза в эритроцитах. Видно, что только три реакции (1, 3 и 10) протекают с высоким изменением свободной энергии, причем равновесие сильно смещено в сторону образования конечных продуктов. Другие реакции легко обратимы. Они могут идти в противоположном направлении при биосинтезе глюкозы (глюконеогенезе), причем с участием тех же ферментов, что и при деградации глюкозы. Для необратимых стадий 1, 3 и 10 в глюконеогенезе используются обходные пути.
Гексозомонофосфатный путь Гексозомонофосфатный путь [ГМП (HMW), часто называемый также пентозофосфатным путем] является окислительным обменом веществ в цитоплазме, в котором, как и в гликолизе, исходным субстратом служит глюкозо-6-фосфат. ГМП поставляет два важных исходных соединения для анаболических процессов: НАДФН + Н+ (NADPH + Н+), необходимый для биосинтеза жирных кислот и изопреноидов и рибозо-5-фосфат, предшественник в биосинтезе нуклеотидов. В процессе окисления глюкозо-6-фосфат превращается в рибулозо-5-фосфат. При этом образуются 1 молекула CO2 и 2 НАДФН + Н+. Значительно более сложная часть пути — восстановительная (Б) — в зависимости от обмена веществ либо превращает часть образованного пентозофосфата снова в гексозофосфат, либо включает его в гликолиз для деградации. В большинстве клеток за счет ГМП разрушается не более 10% глюкозо-6-фосфата. Реакции
Восстановительная часть ГМП показана здесь только схематически. Функция восстановительной ветви состоит в том, чтобы производство НАДФН + Н+ и пентозофосфатов соответствовало метаболическим потребностям клеток. Обычно потребность в НАДФН + Н+ намного выше, чем в пентозофосфатах. В этих условиях 6 молекул рибулозо-5-фосфата под действием трансальдолаз и транскетолаз образуют 5 молекул фруктозо-6-фосфата, которые изомеризуются в 5 молекул глюкозо-6-фосфата. Глюкозо-6-фосфат вновь участвует в окислительной части ГМП в процессе получения НАДФН + Н+. Неоднократное повторение этих реакций позволяет окислить глюкозо-6-фосфат до 6 молекул CO2. При этом образуется 12 молекул НАДФН + Н+, а пентозофосфат не образуется. При взаимном превращении фосфатов сахаров в восстановительной части ГМП особенно важны два фермента. Трансальдолаза [5] переносит С3-звенья от седогептулозо-7-фосфата, кетосахара с 7 атомами углерода, на альдегидную группу глицеральдегид-3-фосфата. Аналогичным образом транскетолаза [4] катализирует перенос С2-фрагмента с одного фосфата сахара на другой. Реакции восстановительной части ГМП обратимы, т.е. гексозофосфаты могут непосредственно превращаться в пентозофосфаты. Это превращение может происходить при высокой потребности клетки в пентозофосфатах, например на стадии репликации ДНК и S-фазе клеточного цикла. Глюконеогенез Некоторые ткани, такие, как мозг и эритроциты, зависят от постоянного снабжения глюкозой. Если получаемое с пищей количество углеводов недостаточно, необходимая концентрация глюкозы в крови может поддерживаться некоторое время за счет расщепления гликогена печенью (см. с. 158). Если истощены также и эти запасы, в печени запускается синтез глюкозы de novo, глюконеогенез (см. с. 302). Наряду с печенью высокой глюконеогенезной активностью обладают также клетки почечных канальцев. Исходными соединениями в глюконеогенезе являются аминокислоты мышечной ткани. При длительном голодании это приводит к массивному распаду мышечного белка. Другими важными исходными веществами для синтеза глюкозы служат лактат, образующийся в эритроцитах и мышечной ткани при недостатке О2, а также глицерин, образующийся при расщеплении жиров. Напротив, жирные кислоты не могут трансформироваться в глюкозу в организме животных, так как в данном случае деградация жирных кислот не является анаплеротическим процессом. В организме человека за счет глюконеогенеза образуется несколько сотен граммов глюкозы в сутки. Многие реакции глюконеогенеза катализируются теми же ферментами, что и процессы гликолиза. Некоторые ферменты специфичны для глюконеогенеза и синтезируются только по мере необходимости под воздействием кортизола и гпюкагона. На схеме представлена только эта группа ферментов. В то время как гликолиз протекает в цитоплазме, глюконеогенез происходит также в митохондриях и эндоплазматическом ретикулуме. Первые стадии реакционной цепи протекают в митохондриях. Причиной такого «обходного» пути является неблагоприятная константа равновесия пируваткиназной реакции. Для перевода пирувата непосредственно в фосфоенолпируват(PEP) недостаточно энергии расщепления АТФ. Пируват, образующийся из лактата или аминокислот, переносится в матрикс митохондрий и там карбоксилируется в оксалоацетат в биотинзависимой реакции, катализируемой пируваткарбоксилазой [2]. Оксалоацетат является промежуточным метаболитом цитратного цикла. Поэтому аминокислоты, которые включаются в цитратный цикл или конвертируются в пируват, могут непосредственно превращаться в глюкозу. Оксалоацетат, образующийся в митохондриальном матриксе, восстанавливается в малат [3]. который может переноситься в цитоплазму с помощью специальных переносчиков. Оксалоацетат может также переноситься из митохондрии в цитоплазму после переаминирования в аспартат. В цитоплазме малат вновь превращается цитоплазматической малатдегидрогеназой в оксалоацетат, который в реакции, катализируемой ГТФ-зависимой РЕР-карбоксикиназой [4], переводится в фосфоенолпируват. Последующие стадии до фруктозо-1,6-дифосфата представляют собой модификации соответствующих реакций гликолиза. При этом для образования 1,3-дифосфоглицерата дополнительно расходуется АТФ. Две глюконеогенез-специфичные фосфатазы отщепляют по очереди фосфатные остатки от фруктозо-1,6-дифосфата. Промежуточной стадией является изомеризация фруктозо-6-фосфата в глюкозо-6-фосфат, одна из реакций гликолиза. Глюкозо-6-фосфатаза печени [5] является мембранным ферментом, локализованным внутри гладкого эндоплазматического ретикулума. Перенос глюкозо-6-фосфата в эндоплазматический ретикулум и возврат образующейся глюкозы в цитоплазму осуществляется специфическими переносчиками. Из цитоплазмы глюкоза поступает в кровь. Глицерин прежде всего фосфорилируется [7] в положении 3. Образующийся 3-глицерофосфат окисляется НАД+-зависимой дегидрогеназой [8] в дигидроксиацетон-3-фосфат, который далее включается в глюконеогенез. Метаболизм гликогена Гликоген служит в животном организме резервом углеводов, из которого по мере метаболической потребности могут высвобождаться глюкозофосфат или глюкоза. Хранение в организме собственно глюкозы неприемлемо из-за ее высокой растворимости: высокие концентрации глюкозы создают в клетке высоко гипертоническую среду, что приводит к притоку воды. Напротив, нерастворимый гликоген осмотически почти неактивен. Гликоген животных, как и амилопектин растений, представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены α(1→4)-гликозидной связью. Связи в точках ветвления находятся в положении α(1→6) примерно каждого 10-го остатка. Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются или удлиняются (при высоком содержании глюкозы) только невосстанавливающие концы древовидной структуры. Удлинение цепи катализируется гликоген-синтазой [2]. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом [УТФ (UTP)] образуется активированный предшественник — УДФ-глюкоза([1]. После этого остаток глюкозы легко переносится с этого промежуточного соединения на гликоген. Когда растущая цепь достигает определенной длины (>11 остатков), специальный фермент ветвления гликогена (1,4→1,6-трансгликозидаза) [3] катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи] Дальнейшее удлинение этого фрагмента осуществляется гликоген-синтазой, образующей α(1→4)-связи. Разветвленная структура гликогена облегчает быстрое освобождение углеводных остатков. Наиболее важным ферментом деградации гликогена является гликоген-фосфорилаза [4], отщепляющая от невосстанавливающего конца цепи остатки глюкозы в виде глюкозо-1-фосфата. Чем больше таких концов, тем больше молекул фосфорилазы могут действовать одновременно. Образование глюкозо-1-фосфата вместо глюкозы имеет то преимущество, что для включения освобожденных остатков глюкозы в гликолиз или ГМП не требуется АТФ. Благодаря структуре гликоген-фосфорилазы, процесс последовательного отщепления останавливается за 4 остатка глюкозы от точки разветвления. Точки ветвления удаляются двумя другими ферментами [5 и 6]. Вначале трисахарид боковой цепи переносится [5] к невосстанавливающему концу главной цепи. Затем 1,6-гликозидаза [6] отщепляет остающийся единичный остаток глюкозы в точке ветвления в виде свободной глюкозы, после чего неразветвленная цепь, может вновь расщепляться фосфорилазой. Баланс гликогена В организме человека может содержаться до 450 г гликогена, треть из которого накапливается в печени, а остальное — главным образом в мышцах. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови в фазе пострезорбции. Поэтому содержание гликогена в печени варьирует в широких пределах. При длительном голодании оно падает почти до нуля, после чего начинается снабжение организма глю Сахарный диабет Сахарный диабет (Diabetes mellitus) — широко распространенное заболевание, которое наблюдается при абсолютном или относительном дефиците инсулина. Нехватка этого пептидного гормона отражается главным образом на обмене углеводов и липидов. Сахарный диабет встречается в двух формах. При диабете I типа (инсулинзависимом сахарном диабете) уже в раннем возрасте происходит гибель инсулинсинтезирующих клеток в результате аутоиммунной реакции. Менее тяжелый диабет II типа (инсулиннезависимая форма) обычно проявляется в более пожилом возрасте. Он
ХИМИЯ УГЛЕВОДОВ Впервые термин «углеводы» был предложен профессором Дерптского (ныне Тартуского) университета К.Г. Шмидтом в 1844 г. В то время предполагали, что все углеводы имеют общую формулу Cm(H2O)n, т.е. углерод + вода. Отсюда название «углеводы». Например, глюкоза и фруктоза имеют формулу С(Н2О)6, тростниковый сахар (сахароза) C12(H2O)11, крахмал [С6(Н2О)5]n и т.д. В дальнейшем оказалось, что ряд соединений, по своим свойствам относящихся к классу углеводов, содержат водород и кислород в несколько иной пропорции, чем указано в общей формуле (например, дезоксирибоза С5Н10О4). В 1927 г. Международная комиссия по реформе химической номенклатуры предложила термин «углеводы» заменить термином «глициды», однако старое название «углеводы» укоренилось и является общепризнанным. Химия углеводов занимает одно из ведущих мест в истории развития органической химии. Тростниковый сахар можно считать первым органическим соединением, выделенным в химически чистом виде. Произведенный в 1861 г. A.M. Бутлеровым синтез (вне организма) углеводов из формальдегида явился первым синтезом представителей одного из трех основных классов веществ (белки, липиды, углеводы), входящих в состав живых организмов. Химическая структура простейших углеводов была выяснена в конце XIX в. в результате фундаментальных исследований Э. Фишера. Значительный вклад в изучение углеводов внесли отечественные ученые А.А. Колли, П.П. Шорыгин, Н.К. Кочетков и др. В 20-е годы нынешнего столетия работами английского исследователя У. Хеуорса были заложены основы структурной химии полисахаридов. Со второй половины XX в. происходит стремительное развитие химии и биохимии углеводов, обусловленное их важным биологическим значением. БИОЛОГИЧЕСКАЯ РОЛЬ УГЛЕВОДОВ Углеводы наряду с белками и липидами являются важнейшими химическими соединениями, входящими в состав живых организмов. У человека и животных углеводы выполняют важные функции: энергетическую (главный вид клеточного топлива), структурную (обязательный компонент большинства внутриклеточных структур) и защитную (участие углеводных компонентов иммуноглобулинов в поддержании иммунитета). Углеводы (рибоза, дезоксирибоза) используются для синтеза нуклеиновых кислот, они являются составными компонентами нуклеотидных ко-ферментов, играющих исключительно важную роль в метаболизме живых существ. В последнее время все большее внимание к себе привлекают смешанные биополимеры, содержащие углеводы: гликопептиды и глико-протеины, гликолипиды и липополисахариды, гликолипопротеины и т.д. Эти вещества выполняют в организме сложные и важные функции. С нарушением обмена углеводов тесно связан ряд заболеваний: сахарный диабет, галактоземия, нарушение в системе депо гликогена, нетолерантность к молоку и т.д. Следует отметить, что в организме человека и животного углеводы присутствуют в меньшем количестве (не более 2% от сухой массы тела), чем белки и липиды; в растительных организмах за счет целлюлозы на долю углеводов приходится до 80% от сухой массы, поэтому в целом в биосфере углеводов больше, чем всех других органических соединений вместе взятых.
КЛАССИФИКАЦИЯ УГЛЕВОДОВ Углеводы можно определить как альдегидные или кетонные производные полиатомных (содержащих более одной ОН-группы) спиртов или как соединения, при гидролизе которых образуются эти производные. Согласно принятой в настоящее время классификации, углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды.
МОНОСАХАРИДЫ Моносахариды можно рассматривать как производные многоатомных спиртов, содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная группа находится в конце цепи, то моносахарид представляет собой альдегид и называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой. Простейшие представители моносахаридов – триозы: глицеральдегид и диоксиацетон. При окислении первичной спиртовой группы трехатомного спирта – глицерола – образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).
Стереоизомерия моносахаридов. Все моносахариды содержат асимметричные атомы углерода: альдотриозы – один центр асимметрии, альдо-тетрозы – 2, альдопентозы – 3, альдогексозы – 4 и т.д. Кетозы содержат на один асимметричный атом меньше, чем альдозы с тем же числом углеродных атомов. Следовательно, кетотриоза диоксиацетон не содержит асимметричных атомов углерода. Все остальные моносахариды могут существовать в виде различных стереоизомеров. Общее число стереоизомеров для любого моносахарида выражается формулой N = 2n, где N – число стереоизомеров, а n – число асимметричных атомов углерода. Как отмечалось, глицеральдегид содержит только один асимметричный атом углерода и поэтому может существовать в виде двух различных стереоизомеров. Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у асимметричного атома углерода расположена с правой стороны, принято считать D-глицеральдегидом, а зеркальное отражение – L-глицеральдегидом:
Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 16 стереоизомеров (24), представителем которых является, например, глюкоза. Для альдопентоз и альдотетроз число сте-реоизомеров равно соответственно 23 = 8 и 22 = 4. Все изомеры моносахаридов подразделяются на D- и L-формы (D-и L-конфигурация) по сходству расположения групп атомов у последнего центра асимметрии с расположением групп у D- и L-глицеральдегида. Природные гексозы: глюкоза, фруктоза, манноза и галактоза – принадлежат, как правило, по стереохимической конфигурации к соединениям D-ряда. Известно, что природные моносахариды обладают оптической активностью. Способность вращать плоскость поляризованного луча света – одна из важнейших особенностей веществ (в том числе моносахаридов), молекулы которых имеют асимметричный атом углерода или асимметричны в целом. Свойство вращать плоскость поляризованного луча вправо обозначают знаком плюс (+), а в противоположную сторону – знаком минус (–). Так, D-глицеральдегид вращает плоскость поляризованного луча вправо, т. е. D-глицеральдегид является D(+)-альдотриозой, а L-гли-церальдегид – L(–)-альдотриозой. Явление мутаротации имеет объяснение. Известно, что альдегиды и кетоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:
Реакция образования полуацеталя возможна и в пределах одной молекулы, если это не связано с пространственными ограничениями. По теории А. Байера, внутримолекулярное взаимодействие спиртовой и карбонильной групп наиболее благоприятно, если оно приводит к образованию пяти- или шестичленных циклов. При образовании полуацеталей возникает новый асимметрический центр (для D-глюкозы это С-1). Шестичленные кольца сахаров называют пиранозами, а пятичленные – фуранозами. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с α-модификацией моносахаридов D-ряда полуаце
|
||||
|
|
Последнее изменение этой страницы: 2021-03-09; просмотров: 253; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.27 (0.018 с.) |





 Гликолиз — это катаболический путь обмена веществ в цитоплазме; он, по-видимому, протекает почти во всех организмах и клетках независимо от того, живут они в аэробных или анаэробных условиях. Баланс гликолиза простой: в аэробных условиях молекула глюкозы деградирует до двух молекул пирувата. Кроме того, образуются по две молекулы АТФ и НАДН + H+ (аэробный гликолиз). В анаэробных условиях пируват претерпевает дальнейшие превращения, обеспечивая при этом регенерацию НАД+. При этом образуются продукты брожения, такие, как лактат или этанол (анаэробный гликолиз). В этих условиях гликолиз является единственным способом получения энергии для синтеза АТФ из АДФ и неорганического фосфата.
Гликолиз — это катаболический путь обмена веществ в цитоплазме; он, по-видимому, протекает почти во всех организмах и клетках независимо от того, живут они в аэробных или анаэробных условиях. Баланс гликолиза простой: в аэробных условиях молекула глюкозы деградирует до двух молекул пирувата. Кроме того, образуются по две молекулы АТФ и НАДН + H+ (аэробный гликолиз). В анаэробных условиях пируват претерпевает дальнейшие превращения, обеспечивая при этом регенерацию НАД+. При этом образуются продукты брожения, такие, как лактат или этанол (анаэробный гликолиз). В этих условиях гликолиз является единственным способом получения энергии для синтеза АТФ из АДФ и неорганического фосфата.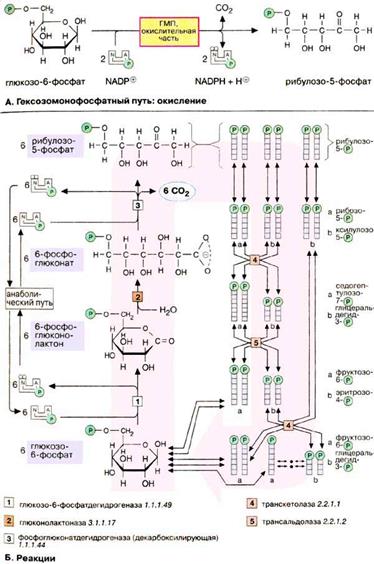 Окислительная часть ГМП начинается с окисления глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой [1]. При этом образуется НАДФН + Н+ и 6-фосфоглюколактон — внутримолекулярный сложный эфир (лактон) 6-фосфоглюконата. Специфическая гидролаза (фермент [2]) расщепляет сложноэфирную связь и оставляет свободной карбоксильную группу 6-фосфоглюконата. Последний фермент окислительной части, фосфоглюконатдегидрогеназа (3), отщепляет карбоксильную группу 6-фосфоглюконата в виде CO2 с одновременным окислением гидроксильной группы при С-3 до кетогруппы. Наряду со второй молекулой НАДФН + Н+ при этом образуется кетопентоза, рибулозо-5-фосфат, которая под действием изомеразы превращается в рибозо-5-фосфат, исходное соединение для нуклеотидного синтеза (на схеме сверху).
Окислительная часть ГМП начинается с окисления глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой [1]. При этом образуется НАДФН + Н+ и 6-фосфоглюколактон — внутримолекулярный сложный эфир (лактон) 6-фосфоглюконата. Специфическая гидролаза (фермент [2]) расщепляет сложноэфирную связь и оставляет свободной карбоксильную группу 6-фосфоглюконата. Последний фермент окислительной части, фосфоглюконатдегидрогеназа (3), отщепляет карбоксильную группу 6-фосфоглюконата в виде CO2 с одновременным окислением гидроксильной группы при С-3 до кетогруппы. Наряду со второй молекулой НАДФН + Н+ при этом образуется кетопентоза, рибулозо-5-фосфат, которая под действием изомеразы превращается в рибозо-5-фосфат, исходное соединение для нуклеотидного синтеза (на схеме сверху).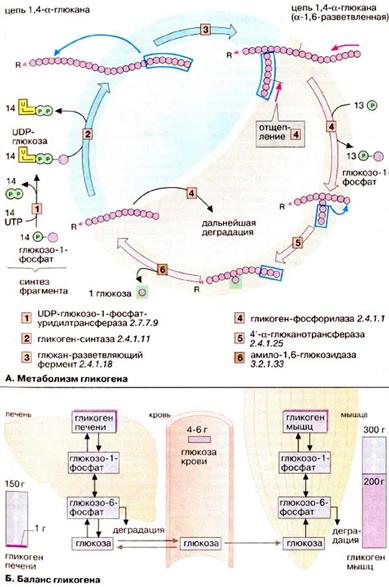 Таким образом, возникает древовидная структура с молекулярной массой >1ּ107 Да (до 50 000 остатков), в которой имеется только одна свободная аномерная ОН-группа, т. е. только один восстанавливающий конец.
Таким образом, возникает древовидная структура с молекулярной массой >1ּ107 Да (до 50 000 остатков), в которой имеется только одна свободная аномерная ОН-группа, т. е. только один восстанавливающий конец. козой с помощью глюконеогенеза. Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. По этой причине колебания содержания гликогена в мышцах меньше, чем в печени.
козой с помощью глюконеогенеза. Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. По этой причине колебания содержания гликогена в мышцах меньше, чем в печени. может быть вызван различными причинами, например пониженной секрецией инсулина или нарушением рецепторных функций.
может быть вызван различными причинами, например пониженной секрецией инсулина или нарушением рецепторных функций.


