Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Вторая афферентная система головного мозга – ретикулярная формация, formatio reticularis
Под этим названием подразумевают совокупность структур, расположенных в центральных отделах мозгового ствола и отличающихся следующими морфологическими особенностями: 1. Нейроны ретикулярной формации имеют отличающее их от других нейронов строение: дендриты их ветвятся очень слабо, нейриты, наоборот, делятся на восходящую и нисходящую ветви, которые отпускают от себя многочисленные коллатерали, благодаря чему аксон может контактировать с огромным числом нервных клеток (при длине в 2 см – с 27 500). 2. Нервные волокна идут в самых различных направлениях, напоминая под микроскопом сеть, что и послужило основанием для Дейтерса назвать ее 120 лет тому назад сетчатой, или ретикулярной, формацией. 3. Клетки ретикулярной формации местами рассеяны, а местами образуют ядра, начало выделению которых положил В. М. Бехтерев, описавший ретикулярное ядро покрышки моста (núcleus reticuláris tegménti póntis). В настоящее время описано 96 отдельных ядер. Область распространения ретикулярной формации точно еще не установлена. На основании физиологических данных, она расположена по всей длине мозгового ствола и занимает центральное положение в продолговатом мозге, мосте, среднем мозге, в гипоталамической области и даже в медиальной части таламусов. Как филогенетически более древняя ретикулярная формация локализуется в покрышке ствола головного мозга. Связи ретикулярной формации. Ретикулярная формация связана со всеми отделами центральной нервной системы. Различают: 1) ретикулопетальные связи, идущие от всех отделов головного мозга; 2) ретикулофугальные связи, идущие к серому веществу и ядрам головного и спинного мозга; 3) ретикулоретикулярные связи (восходящие и нисходящие) между различными ядрами самой ретикулярной формации. Функция. В настоящее время считают, что ретикулярная формация является «генератором энергии» и регулирует процессы, совершающиеся в других отделах центральной нервной системы, включая и кору большого мозга. Особенно важно, что ретикулярная формация оказывает общее (генерализованное) неспецифическое активизирующее воздействие на всю кору головного мозга, (П. К. Анохин), что обеспечивается наличием восходящих про водящих путей от сетчатой формации ко всем долям мозговых полушарий. Поэтому ее называют также восходящей активирующей ретикулярной системой. Будучи связана коллатералями аксонов своих клеток со всеми проходящими через ствол мозга специфическими афферентными проводящими путями, она получает от них импульсы и несет неспецифическую информацию в мозговую кору.
В результате через мозговой ствол проходят в кору мозга две афферентные системы: одна специфическая – это все специфические чувствительные проводящие пути, несущие импульсы от всех рецепторов (экстероинтеро‑ и проприоцепторов) и заканчивающиеся на телах клеток преимущественно IV слоя коры; другая – неспецифическая, образованная ретикулярной формацией и заканчивающаяся на дендритах всех слоев коры. Взаимодействие обеих этих систем обусловливает окончательную реакцию корковых нейронов. Таково современное представление о двух афферентных системах головного мозга. Учитывая столь большое значение ретикулярной формации и ее влияние на кору мозга, некоторые зарубежные исследователи преувеличивают ее роль, считая, что она, располагаясь в центральных частях мозга, составляет особую «центрэнцефалическую» систему, выполняющую функцию сознания и интеграции. Стремление спустить высший уровень интеграции из коры мозга в подкорку не имеет под собой фактических оснований и является антиэволюционным, так как в процессе эволюции наибольшего развития достигает наивысший отдел мозга, т. е. его плащ, а не ствол. Это стремление противоречит материалистической идее нервизма и отражает фрейдизм – идеалистическое учение о ведущей роли не коры, а подкорки. Строение и функция ретикулярной формации полностью еще не раскрыты и составляют предмет дальнейших изысканий.
ЭФФЕРЕНТНЫЕ (НИСХОДЯЩИЕ) ПРОВОДЯЩИЕ ПУТИ Нисходящие двигательные пути идут от коры головного мозга – tráctus corticonucleáris et corticospinális (пирамидная система), от подкорковых ядер переднего мозга – экстрапирамидная система и от мозжечка.
Корково‑спинномозговой (пирамидный) путь, или пирамидная система. Клеточное тело первого нейрона лежит в предцентральной извилине коры большого мозга (гигантские пирамидные клетки). Аксоны этих клеток через coróna radiáta спускаются во внутреннюю капсулу (колено и передние две трети задней ножки), далее в básis pedúnculi cérebri (срединный ее отдел), а затем в pars basiláris моста и продолговатый мозг. Здесь часть волокон пирамидной системы вступает в связь с ядрами черепных нервов. Эта часть пирамидной системы, проходящая через колено внутренней капсулы и связывающая кору большого мозга с ядрами черепных нервов, называется tráctus corticonucleáris [38]. Волокна этого тракта частью переходят на другую сторону, частью остаются на своей стороне. Аксоны клеток, заложенных в ядрах черепных нервов (клеточные тела вторых нейронов), в составе соответственных нервов оканчиваются в скелетной мускулатуре, иннервируемой этими нервами.
Другая часть пирамидной системы, проходящая в передних двух третях задней ножки внутренней капсулы, служит для связи с ядрами спинно‑мозговых нервов, спускается до передних рогов спинного мозга и потому называется tráctus corticospinális. Этот тракт, пройдя в мозговом стволе до продолговатого мозга, образует в нем пирамиды. В последних перекрещивается часть волокон tráctus corticospinális (decussátio pyramídum), которая, спускаясь в спинной мозг, ложится в боковой его канатик, образуя, tráctus corticospinális (pyramidális) laterális . Оставшаяся неперекрещенной часть tráctus corticospinális спускается в переднем канатике спинного мозга, образуя его tráctis corticospinális (pyramidális) antérior (см. рис. 270). Волокна этого пучка постепенно по протяжению спинного мозга также переходят на другую сторону в составе commissúra álba, в результате чего весь tráctus corticospinális оказывается перекрещенным. Благодаря этому кора каждого полушария иннервирует мускулатуру противоположной стороны тела. Двигательные и чувствительные перекресты, происходящие в различных отделах мозга (decussátio pyramídum, commissúra álba, decussátio lemniscórum и др.), представляют, по И. П. Павлову, приспособление нервной системы, направленное на сохранение иннервации при повреждении мозга в каком‑либо месте одной его стороны. Аксоны, составляющие tráctus corticospinális (pyramidális), вступают в связь с двигательными клетками передних рогов спинного мозга, где начинается второе звено[39]. Аксоны лежащих здесь клеток идут в составе передних корешков и далее мышечных нервов к скелетной мускулатуре туловища и конечностей, иннервируемой спинно‑мозговыми нервами. Таким образом, tráctus corticonucleáris и tráctus corticospinális составляют единую пирамидную систему, служащую для сознательного управления скелетной мускулатурой (рис. 353). Эта система особенно развита у человека в связи с прямохождением и сознательным пользованием своим аппаратом движения в процессах труда и членораздельной речью.
Нисходящие пути подкорковых ядер переднего мозга – экстрапирамидная система Пирамидная система, как уже отмечалось выше, начинается в коре большого мозга (V слой, пирамидные клетки). Экстрапирамидная система (рис. 354, 355) слагается из подкорковых образований. В ее состав входят córpus striátum, thalámus, núcleus hypothalámicus postérior, núcleus rúber, substántia nígra и связывающие их проводники белого вещества. Экстрапирамидная система отличается от пирамидной по своему развитию, строению и функции. Она является старейшим в филогенетическом отношении моторно‑тоническим аппаратом, который встречается уже у рыб, у которых имеется еще только бледный шар, pallídum (paleostriátum), у амфибий появляется уже скорлупа, putámen (neostriátum).
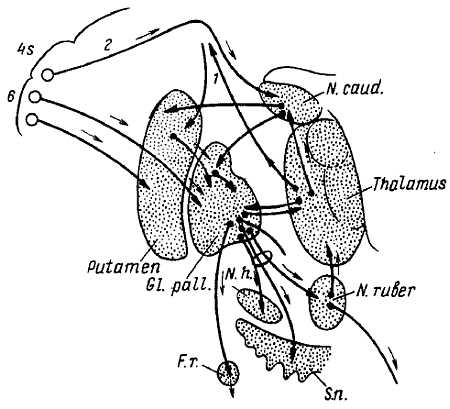
Рис. 354. Связи стриопаллидарной системы и экстрапирамидная система. 6–4s – поля премоторной и двигательной зоны коры мозга; 1 – волокна, восходящие из таламуса в кору; 2 – путь от «тормозных» участков поля 4s в хвостатое ядро (N. caud); Gl. pall . – бледный шар; N. h. – гипоталамическое ядро; N. ruber – красное ядро; S. n. – черная субстанция; F. r. – ретикулярная формация продолговатого мозга. Стрелки указывают направление и «станцию назначения» импульсов.
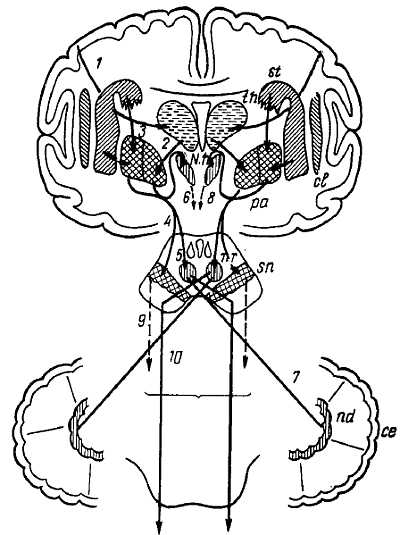
Рис. 355. Схема экстрапирамидной системы. се – кора мозжечка; сl – claustrum; N. h . – гипоталамическое ядро; nd – nucl. dentatus cerebelli; nr – nucl. ruber; pa – pallidum; sn – substantia nigra; st – striatum (nucl. caudatus и putamen); th – thalamus; 1 – tr. corticostrialis; 2 – fibrae thalamopallidales: 3 – fibrae strioppallidales; 4, 5 – связи с substantia nigra и nucl. ruber; 6, 8 – эфферентные волокна гипоталамического ядра; 7 – волокна pedunculus cerebellaris superior; 9 – эфферентные волокна substantia nigra; 10 – tr. rubrospinalis.
На этой стадии развития, когда пирамидная система еще отсутствует, экстрапирамидная система является высшим отделом головного мозга, воспринимающим раздражение от рецепторов органов и посылающим импульсы к мускулатуре через автоматические механизмы спинного мозга. В результате возникают сравнительно простые движения (автоматизированные). У млекопитающих по мере развития переднего мозга и его коры образуется новая кинетическая система – пирамидная, соответствующая новой форме двигательных актов, связанных со все большей специализацией небольших групп мышц. В результате у человека в полной мере развиваются две системы: 1. Пирамидная система – филогенетически более молодая, представлена экранными центрами коры, ведающими сознательными движениями человека. Через пирамидную систему осуществляется также в движениях корковая деятельность, основанная на условных рефлексах. 2. Экстрапирамидная система – филогенетически более старая, состоящая из подкорковых ядер. У человека она играет подчиненную роль и осуществляет высшие безусловные рефлексы, поддерживая тонус мускулатуры и автоматически регулируя ее работу (непроизвольная автоматическая иннервация скелетной мускулатуры). Эта автоматическая регуляция мышц осуществляется благодаря связям всех компонентов экстрапирамидной системы между собой и с núcleus rúber, от которого идет нисходящий двигательный путь к передним рогам серого вещества спинного мозга, tráctus rubrospinális. Этот тракт начинается в клетках красного ядра, переходит через срединную плоскость на уровне верхних холмиков крыши среднего мозга, образуя вентральный перекрест (decussátio ventrális tegménti), и спускается через мозговой ствол в боковые канатики спинного мозга, после чего заканчивается на двигательных нейронах передних рогов серого вещества. Таким образом, экстрапирамидная система действует на спинной мозг через красное ядро, которое составляет важнейшую часть этой системы.
К работе экстрапирамидной системы имеют отношение нисходящие мозжечковые пути, а также ретикулярно‑спинномозговой путь, которому в настоящее время придается большое значение в регуляции двигательной активности спинного мозга.
Нисходящие двигательные пути мозжечка Мозжечок принимает участие в контроле двигательных нейронов спинного мозга (мышечная координация, поддержание равновесия, сохранение мышечного тонуса и преодоление инерции и силы тяжести). Это осуществляется с помощью tráctus cerebellorubrospinális (см. рис. 352). Клеточное тело первого звена этого пути лежит в коре мозжечка (грушевидные нейроциты). Их аксоны заканчиваются в núcleus dentátus cerebélli и, возможно, в других ядрах мозжечка, где начинается второе звено . Аксоны вторых нейронов идут через верхние мозжечковые ножки к среднему мозгу и оканчиваются в núcleus rúЬег. Здесь помещаются клетки третьего звена , аксоны которых в составе tráctus rubrospinális, переключившись в двигательных нейронах передних рогов спинного мозга (четвертое звено ), достигают скелетной мускулатуры.
Нисходящие пути коры большого мозга к мозжечку Кора большого мозга, ведающая всеми процессами организма, держит в своем подчинении и мозжечок как важнейший проприоцептивный центр, связанный с движениями тела. Это достигается наличием специального нисходящего пути от коры большого мозга к коре мозжечка – tráctus corticopontocerebelláris (см. рис. 352). Первое звено этого пути состоит из нейронов, клеточные тела которых заложены в коре большого мозга, а аксоны спускаются к ядрам моста, núclei (próprii) póntis. Эти нейроны составляют отдельные пучки, которые соответственно различным долям мозга называются tráctus frorttopontínus, occipitopontínus, temporopontínus et parietopontínus. В ядрах моста начинаются вторые нейроны , аксоны которых образуют tráctus роntocerebelláris, идущий на противоположную сторону моста, и в составе средних мозжечковых ножек он достигает коры полушарий мозжечка (neocerebellum).
Таким образом, устанавливается связь между корой большого мозга и полушариями мозжечка. (Полушария головного мозга связаны с противоположными полушариями мозжечка.) Оба эти отдела головного мозга являются более молодыми и в своем развитии взаимосвязаны. Чем сильнее развиты кора и полушария большого мозга, тем сильнее развиты кора и полушария мозжечка. Так как связь этих отделов головного мозга осуществляется через мост, то и степень развития последнего определяется развитием мозговой коры. Следовательно, три пары ножек мозжечка обеспечивают его многосторонние связи: через нижние ножки он получает импульсы из спинного мозга и продолговатого мозга, через средние – из коры полушарий большого мозга; в составе верхних ножек проходит главный эфферентный путь мозжечка, по которому мозжечковые импульсы передаются на клетки передних рогов спинного мозга. Связь полушарий головного мозга с полушариями мозжечка, т. е. с его новой частью (neocerebéllum), перекрестная, связь же червя, т. е. старой части мозжечка (paleocerebéllum), со спинным мозгом главным образом прямая, гомолатеральная.
УЧЕНИЕ ОБ ОРГАНАХ ЧУВСТВ (ЭСТЕЗИОЛОГИЯ). ÓRGANA SÉNSUUMO
Общие данные
Органами чувств, или анализаторами, называются приборы, посредством которых нервная система получает раздражения от внешней среды, а также от органов самого тела и воспринимает эти раздражения в виде ощущений. Показания органов чувств являются источниками представлений об окружающем нас мире. «Иначе, как через ощущения, мы ни о каких формах вещества и ни о каких формах движения ничего узнать не можем…» (Ленин В. И. Полн. собр. соч., т. 18, с. 320). Поэтому В. И. Ленин считал физиологию органов чувств одной из наук, лежащих в основе построения диалектико‑материалистической теории познания. Процесс чувственного познания совершается у человека по шести каналам: осязание, слух, зрение, вкус, обоняние, земное тяготение. Шесть органов чувств дают человеку многообразную информацию об окружающем объективном мире, которая отражается в сознании в виде субъективных образов – ощущений, восприятий и представлений памяти. Живая протоплазма обладает раздражимостью и способностью отвечать на раздражение. В процессе филогенеза эта способность особенно развивается у специализированных клеток покровного эпителия под влиянием внешних раздражений и клеток кишечного эпителия под влиянием раздражения пищей. Специализированные клетки эпителия уже у кишечнополостных оказываются связанными с нервной системой. В некоторых участках тела, например на щупальцах, в области рта, специализированные клетки, обладающие повышенной возбудимостью, образуют скопления, из которых возникают простейшие органы чувств. В дальнейшем в зависимости от положения этих клеток происходит их специализация по отношению к раздражителям. Так, клетки ротовой области специализируются к восприятию химических раздражений (обоняние, вкус), клетки на выступающих частях тела – к восприятию механических раздражений (осязание) и т. д. Развитие органов чувств обусловлено значением их для приспособления к условиям существования. Например, собака тонко воспринимает запах ничтожных концентраций органических кислот, выделяемых телом животных (запах следов), и плохо разбирается в запахе растений, которые не имеют для нее биологического значения. Возрастание тонкости анализа внешнего мира обусловлено не только усложнением строения и функции органов чувств, но прежде всего усложнением нервной системы. Особенное значение для анализа внешнего мира приобретает развитие головного мозга (особенно его коры), отчего Ф. Энгельс называет органы чувств «орудиями мозга». Возникающие в силу тех или иных раздражений нервные возбуждения воспринимаются нами в форме различных ощущений. Как учит ленинская теория отражения, ощущение – это отражение в сознании человека предметов и явлений внешнего мира в результате их воздействия на органы чувств. Так, например, световая энергия, действуя на сетчатку глаза, вызывает нервные импульсы, которые, передаваясь по нервной системе, вызывают в нашем сознании зрительные ощущения. «…Ощущение… есть превращение энергии внешнего раздражения в факт сознания» (Ленин В. И. Пол. собр. соч., т. 18, с. 46). Для возникновения ощущений необходимы: приборы, воспринимающие раздражение, нервы, по которым передается это раздражение, и мозг, где оно превращается в факт сознания. Весь этот аппарат, необходимый для возникновения ощущения, И. П. Павлов назвал анализатором (см. также «Морфологические основы динамической локализации функций…»). «Анализатор – это такой прибор, который имеет своей задачей разлагать сложность внешнего мира на отдельные элементы» (Павлов И. П. Лекции по физиологии, 1952, с. 445). Каждый анализатор состоит из трех частей: 1) рецептор – трансформатор энергии раздражения в нервный процесс; 2) кондуктор – проводник нервного возбуждения и 3) корковый конец анализатора, где возбуждение воспринимается как ощущение. Различают две группы ощущений: 1. Ощущения, отражающие свойства предметов и явлений окружающего материального мира: осязание, т. е. ощущение прикосновения и давления, температурное чувство (тепла, холода) и боль; затем ощущения слуховые, зрительные, вкусовые, обонятельные и земного притяжения. 2. Ощущения, отражающие движения отдельных частей тела и состояние внутренних органов (двигательные ощущения, ощущение равновесия тела, ощущения органов). Соответственно этому все органы чувств делят на две группы: 1. Органы внешних чувств, получающие нервные импульсы из экстероцептавного поля, – экстероцепторы . Их шесть: органы кожного чувства, чувства земного тяготения (гравитации), слуха, зрения, вкуса и обоняния. 2. Органы внутренних ощущений: а) получающие импульсы из проприоцептивного поля (мышечно‑суставное чувство, тесно связанное с чувством земного притяжения) – проприоцепторы ; б) органы, воспринимающие нервные импульсы из интероцептивного поля (внутренностей и сосудов), – интероцепторы . Ощущения, идущие из внутренних органов, обычно неопределенны и при нормальном состоянии этих органов не достигают сознания, сказываясь только «общим самочувствием». Вообще все внутренние процессы, регулируемые вегетативной нервной системой, протекают без нашего ведома и только при болезненных расстройствах дают о себе знать обычно более или менее сильной болью. Подробно об интероцептивном анализаторе было изложено в специальной главе. Из возбуждений, идущих от проприоцептивного поля, надо упомянуть только мышечно‑суставное чувство, благодаря которому воспринимается ощущение положения частей тела и происходит координация движений. С одной стороны, это чувство комбинируется с кожной чувствительностью (чувство стереогноза), а с другой, стоит в связи с органом гравитации, дающим ориентацию по отношению к гравитационному полю, который может быть рассмотрен также как статокинетический аппарат, обеспечивающий равновесие тела. Нервные окончания (в мышцах, костях, сухожилиях и суставах) и проводники мышечно‑суставного чувства были описаны при изложении двигательного анализатора. В данном разделе будут рассмотрены только органы, воспринимающие ощущения, получаемые из внешнего мира, – экстероцепторы. Общий план воспринимающих приборов у всех классов животных более или менее одинаков, несмотря на последующие значительные усложнения в деталях. Основным элементом, за исключением органов кожного чувства, у наземных животных являются особые чувствительные клетки, которые в процессе развития всегда происходят из эпителия наружного листка (эктодермы), который уже по своему положению находится в соприкосновении с окружающим миром. Каждая такая клетка на одном конце, обращенном к наружной поверхности, несет штифтик или воспринимающие волоски, а с другой стороны отдает (в органе обоняния и зрения) отросток, идущий на соединение с отростками нервных клеток проводящих нейронов. В других органах (вкуса и слуха) чувствительная клетка, не давая центрального отростка, оплетается концевыми разветвлениями подходящего к ней афферентного нерва. Первый тип чувствительных клеток сравнительно со вторым видом нужно считать первичным. У водных животных такая форма воспринимающих элементов встречается и в кожных покровах, где эти элементы подвергаются увлажнению окружающей жидкостью. В коже наземных животных чувствительных клеток не бывает, и рецепторные нервные волокна оканчиваются или свободно между клетками эпителиального покрова, или же имеют на своих концах особого рода концевые тельца. В образовании органов чувств принимает также участие мезодерма, но только вторичным порядком, образуя для них защитные, поддерживающие и вспомогательные приспособления. Эти приспособления, обрастающие и дополняющие чувствительные клетки, т. е. рецепторы, образуют вместе с ними периферические отделы органов чувств: кожа, ухо, глаз, язык, нос. Например, зрительным рецептором являются чувствительные клетки сетчатки (палочки и колбочки), а периферическим отделом – весь глаз. Кроме деления органов чувств на 2 группы, все анализаторы можно классифицировать с точки зрения учения И. П. Павлова о двух сигнальных системах следующим образом: I. Анализаторы первой сигнальной системы (конкретно‑наглядное мышление): А. Анализаторы внешнего мира – экстероцепторы (органы кожного чувства, слуха, зрения, вкуса, обоняния и гравитации). Б. Анализаторы внутреннего мира организма: 1. Проприоцепторы, несущие раздражение от органов животной жизни (мышечно‑суставное чувство). 2. Интероцепторы, несущие раздражение от органов растительной жизни (внутренности, сосуды). II. Анализаторы второй сигнальной системы (абстрактно‑логическое мышление): 1. Анализаторы устной речи. 2. Анализаторы письменной речи. Анализаторы первой и второй сигнальных систем имеют резкие анатомические отличия. Анализаторы первой сигнальной системы обладают каждый всеми тремя компонентами (рецептор, кондуктор и корковый конец). Анализаторы второй сигнальной системы лишены своих рецепторов и кондукторов, а имеют только корковые концы (корковые концы речевых анализаторов); они воспринимают свои сигналы (вторые сигналы) на базе первых сигналов, составляющих первую сигнальную систему, без которой они не функционируют. Этим подчеркивается и разделение, и объединение корковых концов всех анализаторов, составляющих единую кору большого мозга, где совершается «…превращение энергии внешнего раздражения в факт сознания» (Ленин В. И. Полн. собр. соч., т. 18, с. 46).
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 89; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.38.125 (0.056 с.) |