Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Вентиляция и перфузия кровью легких
10.2.1. Вентиляция легких Вентиляцией легких обозначают процесс обмена воздуха между легкими и атмосферой. Количественным показателем вентиляции легких служит минутный объем дыхания, определяемый как количество воздуха, которое проходит (или вентилируется) через легкие в 1 мин. В покое у человека минутный объем дыхания составляет 6—8 л/мин. Только часть воздуха, которым вентилируются легкие, достигает альвеолярного пространства и непосредственно участвует в газообмене с кровью. Эта часть вентиляции легких называется альвеолярной вентиляцией. В покое альвеолярная вентиляция равна в среднем 3,5—4,5 л/мин. Основная функция альвеолярной вентиляции заключается в поддержании необходимой для газообмена концентрации О2 и СО2 в воздухе альвеол. Легкие состоят из воздухопроводящей (дыхательные пути) и респираторной зон (альвеолы). Дыхательные пути, начиная от трахеи и до альвеол, делятся по типу дихотомии и образуют 23 генерации элементов дыхательного тракта (рис. 10.11). В воздухопроводящей или кондуктивной зонах легких (16 генераций) отсутствует газообмен между воздухом и кровью, поскольку в этих отделах дыхательные пути не имеют достаточной для этого процесса сосудистой сети, а стенки дыхательных путей, из-за их значительной толщины, препятствуют обмену газов через них. Этот отдел воздухоносных путей называется анатомическим мертвым пространством, объем которого составляет в среднем 175 мл. На рис. 10.12 показано, каким образом воздух, заполняющий анатомическое мертвое пространство в конце выдоха, смешивается с «полезным», т. е. атмосферным воздухом и вновь поступает в альвеолярное пространство легких. Дыхательные бронхиолы 17—19-й генераций относят к переходной (транзиторной) зоне, в которой начинается газообмен в малочисленных альвеолах (2 % от общего числа альвеол). Альвеолярные ходы и альвеолярные мешочки, непосредственно переходящие в альвеолы, образуют альвеолярное пространство, в области которого происходит в легких газообмен О2 и СО2 с кровью. Однако у здоровых людей и, особенно, у пациентов с
Рис. 10.11. Схема дыхательных путей легких человека. Дыхательные пути от уровня трахеи (1-я генерация) до долевых бронхов (2—4-я генерации деления) поддерживают свой просвет благодаря хрящевым кольцам в их стенке. Дыхательные пути от сегментарных бронхов (5—11-я генерации) до терминальных бронхиол (12—16-я генерации) стабилизируют свой просвет с помощью тонуса гладких мышц их стенок. 1—16-я генерации дыхательных путей образуют воздухопроводящую зону легких, в которой не происходит газообмена. Респираторная зона легких имеет длину порядка 5 мм и включает первичные дольки или ацинусы: дыхательные бронхиолы (17—19-я генерации) и альвеолярные протоки (20—22-я генерации). Альвеолярные мешочки состоят из многочисленных альвеол (23-я генерация), альвеолярная мембрана которых является идеальным местом для диффузии О2 и СО2.
заболеваниями легких часть альвеолярного пространства может вентилироваться, но при этом не участвовать в газообмене, поскольку эти отделы легких не перфузируются кровью. Сумму объемов таких областей легких и анатомического мертвого пространства обозначают как физиологическое мертвое пространство. Увеличение физиологического мертвого пространства в легких приводит к недостаточному снабжению тканей организма кислородом и к увеличению содержания в крови углекислого газа, что нарушает в ней газовый гомеостазис.
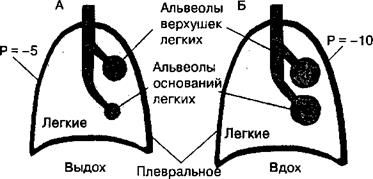
10.2.2. Перфузия легких кровью Эффективность выполнения основной функции дыхательной системы зависит от соответствия перфузии (кровотока) в регионах легких с их вентиляцией. Так, хороший кровоток будет недостаточным для газообмена в регионах легких, если они слабо вентилируются воздухом с низким содержанием кислорода, при этом незначительный объем вентиляции легких не позволит удалить из крови углекислый газ. Соответственно, при слабом кровотоке в регионах легких возрастает объем функционального мертвого пространства, и перфузия легких кровью будет недостаточной для транспорта в ней нормального количества газов. Наконец, перфузия кровью регионов легких с недостаточной вентиляцией этих же отделов называется шунтированием, и это состояние является неадекватным для нормального газообмена. При шунтировании венозная кровь в легких не обогащается кислородом, что снижает его содержание в крови организма. В нормальных физиологических условиях фактор гравитации оказывает наиболее выраженный эффект на вентиляцию и перфузию регионов легких кровью. 10.2.3. Эффект гравитации на вентиляцию и перфузию легких кровью Легкие окружены плевральным пространством, отрицательное давление в котором изменяется от “5 до —10 см водн. ст. в различные фазы дыхательного цикла. Этот фактор взаимодействует с эффектом гравитации на жидкие среды, прежде всего кровь, содержащуюся в артериальных и венозных сосудах тканей легких. В результате под действием силы тяжести на ткань легких величина внутриплеврального давления на уровне основания легких у человека в положении стоя менее отрицательная относительно атмосферного, чем в области верхушек легкого. Поэтому альвеолы верхушек легких имеют большие размеры, а стенка их растянута и более напряжена, чем у альвеол нижних участков легких. Альвеолы на уровне основания легких растянуты в незначительной степени и имеют значительно больше потенциальные возможности для растягивания и вентиляции, чем в области верхушек. Поэтому растянутые альвеолы верхушки легких вентилируются меньше, чем альвеолы основания (рис. 10.13). Эти различия в вентиляции отделов легких приводят к тому, что вдыхаемый воздух неравномерно распределяется в отделах альвеолярного пространства. Особенности распределения воздуха, вдыхаемого в легкие, дополняется разницей в величине кровотока на уровне верхушек и основания легких. Относительно положения тела в пространстве кровоток в верхних и нижних отделах легкого различается под влиянием фактора гравитации. У человека в вертикальном положении тела величина легочного кровотока на единицу объема ткани легкого линейно убывает в направлении
пространство Рис. 10.13. Влияние внутриплеврального давления и фактора гравитации на размеры альвеол верхних и нижних отделов легких. Между верхушками и основанием легких имеется градиент внутриплеврального давления, возникающий под влиянием гравитации на массу жидких сред и ткань легких.
В результате размеры альвеол в верхушках легких больше, чем в основании (А). Альвеолы в нижних отделах легких имеют большие потенциальные возможности для увеличения в них вентиляции легких при вдохе, чем альвеолы в области верхних отделов легких (Б).
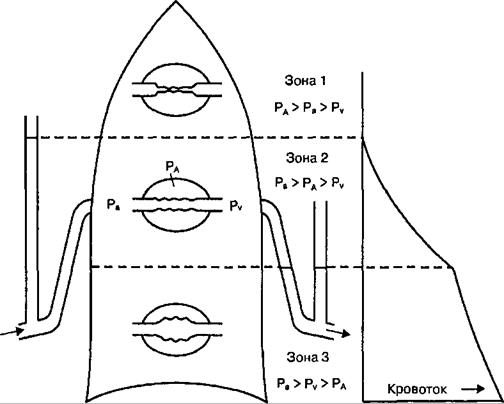
снизу вверх, и меньше всего снабжаются кровью верхушки легких. Соответственно в положении тела человека на спине кровоток в нижних (дорсальных) отделах легких становится выше, чем в верхних (вентральных). Это обусловлено тем, что артериальная кровь, поступающая в легкие из правого желудочка, проходит по сосудам легких из областей низкого внутриплеврального давления в области тонкостенных капилляров, которые окружены альвеолами, содержащими воздух под давлением, близким к атмосферному. Поэтому в зависимости от соотношения давления в альвеолах (РА), мелких артериях (Ра) и мелких легочных венах (Pv) легкие разделены на функциональные зоны Веста (рис. 10.14). В верхушках легких (зона 1) могут возникнуть области с давлением в легочных капиллярах (особенно в фазу диастолы) ниже альвеолярного (Ра > Ра > Pv)- Капилляры в таких зонах могут спадаться, и кровоток через них становится невозможным. Такие участки легких вентилируются, но не участвуют в газообмене и формируют альвеолярное мертвое пространство. В средних отделах легких (зона 2) под действием гравитации давление в альвеолах, как правило, превышает венозное (Ра > РА > Pv)- Поэтому величину кровотока в зоне 2 по Весту определяет разность между артериальным и альвеолярным давлениями. В зоне 2 практически не возникает альвеолярное мертвое пространство. В нижних отделах легких (зона 3) давление в легочных венах выше альвеолярного (Ра > Pv > РА) и величина кровотока, как и в обычных сосудах, определяется разницей между артериальным и венозным давлениями. Величина зон Веста динамично изменяется в зависимости от положения тела в пространстве или глубины дыхания. При выдохе на уровне функциональной остаточной емкости примерно 2/3 объема легких может занимать зона 2. После глубокой экспирации (на уровне остаточного объема) большая часть легких по соотношению перфузии кровью и вентиляции соответствует зоне 3 Веста. Относительная однонаправленность изменения градиента внутриплеврального давления и влияния гравитации на кровоток в легких от верхних отделов легких к нижним теме не менее не сопряжены в каждом отдельном регионе легких. 10.2.3. Коэффициент вентиляционно-перфузионных отношений в легких Коэффициент представляет собой отношение величины вентиляции легких (V) к величине их перфузии кровью (Q). При адекватности вентиляционно-перфузионных отношений (V/Q) величина коэффициента близка к 1. В обычных физиологических условиях в разных регионах легких, как правило, коэффициент варьирует: 1<V/Q >1. Региональные отличия значения этого коэффициента обусловлены действием гравитации либо в случае появления в каком-либо регионе легких феномена шунтирования (рис. 10.15). Однако в целом 97—98 % от общего количества О2 и СО2, участвующих в газообмене в легких, обменивается между альвеолярным пространством и кровью легочных капилляров в условиях полного соответствия величины вентиляции и перфузии легких кровью (V/Q =1).
Газообмен в легких Обмен газов между кровью и воздухом относится к основной функции легких. Воздух, поступающий в легкие при вдохе, нагревается и насыщается водяными парами при движении в дыхательных путях и достигает альвеолярного пространства, имея температуру 37 °C. Парциальное давление
Рис. 10.14. Модель, связывающая неравномерность распределения легочного кровотока при вертикальном расположении тела человека с величиной давления, действующего на капилляры.
В зоне 1 (верхушки легких) альвеолярное давление (РА) превышает давление в артериолах (Ра) и кровоток ограничен. В средней зоне легких (зона 2), где Рй > РА, кровоток больше, чем в зоне 1. В основаниях легких (зона 3) кровоток усилен и определяется разностью давления в артериолах (Ра) и венулах (Pv). В центре схемы легкого — легочные капилляры; вертикальные трубочки по сторонам легкого — манометры.
При прекращении вентиляции в каком-либо регионе легких увеличивается их функциональное мертвое пространство (а). При этом венозная кровь перфузирует этот отдел легких и, не обогащаясь кислородом, поступает в большой круг кровообращения. Нормальное вентиляционно-перфузионное отношение формируется, когда вентиляция регионов легких соответствует величине их перфузии кровью (б). При отсутствии кровотока в каком-либо регионе легких (в) вентиляция также не обеспечивает нормальное вентиляционно-перфузионное отношение. V — вентиляция легких, Q — кровоток в легких. водяных паров в альвеолярном воздухе при этой температуре составляет 47 мм рт. ст. Поэтому согласно закону парциальных давлений Дальтона вдыхаемый воздух находится в разведенном водяными парами состоянии и парциальное давление кислорода в нем меньше, чем в атмосферном воздухе. Обмен кислорода и углекислого газа в легких происходит в результате разницы парциального давления этих газов в воздухе альвеолярного пространства и их напряжения в крови легочных капилляров. Процесс движения газа из области высокой концентрации в область с низкой его концентрацией обусловлен диффузией. Кровь легочных капилляров отделена от воздуха, заполняющего альвеолы, альвеолярной мембраной, через которую газообмен происходит путем пассивной диффузии. Процесс перехода газов между альвеолярным пространством и кровью легких объясняется диффузионной теорией.
10.3.1. Состав альвеолярного воздуха Газовый состав альвеолярного воздуха обусловлен альвеолярной вентиляцией и скоростью диффузии О2 и СО2 через альвеолярную мембрану. В обычных условиях у человека количество О2, поступающего в единицу времени в альвеолы из атмосферного воздуха, равно количеству О2, диффундирующего из альвеол в кровь легочных капилляров. Равным образом количество СО2, поступающего в альвеолы из венозной крови, равно количеству СО2, которое выводится из альвеол в атмосферу. Поэтому в норме парциальное давление О2 и СО2 в альвеолярном воздухе остается практически постоянным, что поддерживает процесс газообмена между альвеолярным воздухом и кровью капилляров легких. Газовый состав альвеолярного воздуха отличается от атмосферного воздуха тем, что в нем Таблица 10.1. Парциальное давление газов в воздушной среде легких
меньше процентное содержание кислорода и выше процент углекислого газа. Состав альвеолярного воздуха отличается от выдыхаемого воздуха большим содержанием углекислого газа и меньшим содержанием кислорода (табл. 10.1). 10.3.2, Напряжение газов в крови капилляров легких Диффузия газов через альвеолярную мембрану происходит между альвеолярным воздухом и венозной, а также артериальной кровью легочных капилляров. В табл. 10.2 приведены стандартные величины напряжения дыхательных газов в артериальной и венозной крови легочных капилляров. Градиенты парциального давления кислорода и углекислого газа обусловливают процесс пассивной диффузии через альвеолярную мембрану кислорода из альвеол в венозную кровь (градиент 60 мм рт. ст.), а углекислого газа — из венозной крови в альвеолы (градиент 6 мм рт. ст.). Парциальное давление азота по обе стороны альвеолярной мембраны остается постоянным, поскольку этот газ не потребляется и не продуцируется тканями организма. При этом сумма парциального давления всех газов, растворенных в тканях организма, меньше, чем величина атмосферного давления, благодаря чему газы в тканях не находятся в газообразной форме. Если величина атмосферного давления будет меньше, чем парциальное давление газов в тканях и в крови, то газы начинают выделяться из крови в виде пузырьков, вызывая тяжелые нарушения в кровоснабжении тканей организма (кессонная болезнь). 10.3.3. Скорость диффузии О2 и СО2 в легких Скорость диффузии (M/t) кислорода и углекислого газа через альвеолярную мембрану количественно характеризуется законом диффузии Фика. Согласно этому закону газообмен (M/t) в легких прямо пропорционален градиенту (ДР) концентрации О2 и СО2 по обе стороны от альвеолярной мембраны, площади ее поверхности (S), коэффициентам (к) растворимо- Таблица 10.2. Напряжение дыхательных газов в артериальной и венозной крови легочных капилляров
Альвеолярное Альвеолярная Просвет
Рис. 10.16. Диффузия газов через альвеолярную мембрану. Диффузия газов в легких осуществляется по градиентам концентрации О2 и СО2 между альвеолярным пространством и кровью капилляров легких, которые разделены альвеолярной мембраной. При этом диффузия тем эффективнее, чем тоньше альвеолярная мембрана и области контакта альвеолоцитов и эндотелиоцитов. Поэтому альвеолярная мембрана образована уплощенными частями альвеолоцитов I порядка (0,2 мкм) и эндотелиоцитов капилляров легких (0, 2 мкм), между которыми находится тонкая общая базальная мембрана (0,1 мкм) этих клеток. В состав мембраны входит также моно- молекулярный слой сурфактант а. Мембрана эритроцитов является препятствием для диффузии газов в легких. сти О2 и СО2 в биологических средах альвеолярной мембраны и обратно пропорционален толщине альвеолярной мембраны (L), а также молекулярной массе газов (М). Формула этой зависимости имеет следующий вид: М ДР • S • к * L • JM Структура легких образует максимальное по величине поле для диффузии газов через альвеолярную стенку, которая имеет минимальную толщину (рис. 10.16). Так, количество альвеол в одном легком человека приблизительно равно 300 млн. Суммарная площадь альвеолярной мембраны, через которую происходит обмен газов между альвеолярным воздухом и венозной кровью, имеет огромные размеры (порядка 100 м2), а толщина альвеолярной мембраны составляет лишь — 0,3—2,0 мкм. В обычных условиях диффузия газов через альвеолярную мембрану происходит в течение очень короткого отрезка времени (не более 3/4 с), пока кровь проходит через капилляры легких. Даже при физической работе, когда эритроциты проходят капилляры легкого в среднем за т/4 с, указанные выше структурные особенности альвеолярной мембраны создают оптимальные условия для формирования равновесия парциальных давлений О2 и СО2 между альвеолярным воздухом и кровью капилляров легких (рис. 10.17). В уравнении Фика константы диффузии (к) пропорциональны растворимости газа в альвеолярной мембране. Углекислый газ имеет примерно в 20 раз большую растворимость в альвеолярной мембране, чем кислород. Поэтому, несмотря на существенное различие в градиентах парциальных давлений О2 и СО2 по обе стороны от альвеолярной мембраны, Альвеолярный газ (РО2 = 106 мм рт.ст.)
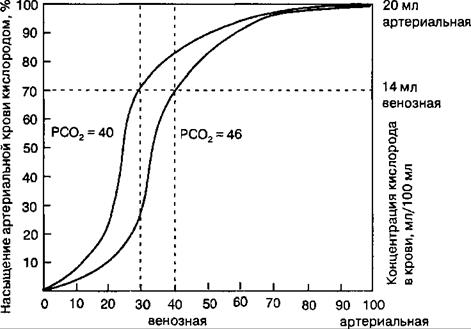
Рис. 10.17. Градиенты парциального давления дыхательных газов в смешанной венозной крови легочной артерии, альвеолярном воздухе и артериальной крови. Равновесие парциальных давлений углекислого газа и кислорода между альвеолярным воздухом и кровью легочных капилляров достигается в течение короткого времени (1/4—3Л с) движения плазмы крови и эритроцитов в капиллярах легких. диффузия этих газов совершается за очень короткий отрезок времени движения эритроцитов крови через легочные капилляры. Газообмен через альвеолярную мембрану количественно оценивается диффузионной способностью легких, которая измеряется количеством газа (мл), проходящего через эту мембрану за 1 мин при разнице давления газа по обе стороны мембраны в 1 мм рт. ст. Наибольшее сопротивление диффузии О2 в легких создают альвеолярная мембрана и мембрана эритроцитов, в меньшей степени — плазма крови в капиллярах. У взрослого человека в покое диффузионная способность легких О2 равна 20—25 мл • мин"*1 • мм рт. ст."1. СО2, как полярная молекула (О=С=О), диффундирует через указанные мембраны чрезвычайно быстро, благодаря высокой растворимости этого газа в альвеолярной мембране. Диффузионная способность легких СО2 равна 400—450 мл • мин"1 • мм рт. ст.-1. Транспорт газов кровью Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении. 10.4.1. Транспорт кислорода При 37 °C растворимость О2 в жидкости составляет 0,225 мл-л"1-кПа"1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл О2, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах. Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется оксигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при РО2 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения ки-
Парциальное давление кислорода, мм рт. ст. Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РСО2 = 40 мм рт. ст. (артериальная кровь) и РСО2 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена). слородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием РО2 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду. Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях градиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м. Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток. Один грамм гемоглобина связывает 1,34 мл О2. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл О2. Количество О2, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание О2 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл О2. 10.4.1.1. Изменение сродства гемоглобина к кислороду Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов. Основной фактор — это связывание кислорода с геминовой группой молекулы гемоглобина. При этом чем больше геминовых групп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком РО2, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком РО2 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода. Когда эритроциты, содержащие гемоглобин, достигают тканей, то кислород из эритроцитов диффундирует в клетки. В мышцах он поступает в своеобразного депо кислорода — в молекулы миоглобина, из которого кислород используется в биологическом окислении мышц. Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена низким РО2 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение кислорода, необходимое для поддержания нормального метаболизма, составляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем диффузии из капилляров достигает метаболически активных клеток. Некоторые ткани приспособлены к низкому содержанию РО2 в капиллярах крови, что компенсируется высокой плотностью капилляров на единицу объема тканей. Например, в скелетной и сердечной мышцах РО2 в капиллярах может снизиться чрезвычайно быстро во время сокращения. В мышечных клетках содержится белок миоглобин, который имеет более высокое сродство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается кислородом и способствует его диффузии из крови в скелетную и сердечную мышцы, где он обусловливает процессы биологического окисления. Эти ткани способны экстрагировать до 70 % кислорода из крови, проходящей через них, что обусловлено снижением сродства гемоглобина к кислороду под влиянием температуры тканей и pH. Эффект pH и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое pH понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и СО2. Если pH плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном pH 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда pH снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 73 больше, чем при постоянном pH. Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани. Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении РО2 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3- ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей. 10.4.2. Транспорт углекислого газа СО2 является продуктом метаболизма клеток тканей и поэтому переносится кровью от тканей к легким. Углекислый газ выполняет жизненно важную роль в поддержании во внутренних средах организма уровня pH механизмами кислотно-основного равновесия. Поэтому транспорт углекислого газа кровью тесно взаимосвязан с этими механизмами (см. глава 15). В плазме крови небольшое количество СО2 находится в растворенном состоянии; при РСО2= 40 мм рт. ст. переносится 2,5 мл/100 мл крови СО2, или 5 %. Количество растворенного в плазме углекислого газа в линейной зависимости возрастает от уровня РСО2. В плазме крови СО2 реагирует с водой с образованием Н+ и НСО^. Увеличение напряжения углекислого газа в плазме крови вызывает уменьшение величины ее pH. Напряжение СО2 в плазме крови может быть изменено функцией внешнего дыхания, а количество ионов водорода или pH — буферными системами крови и НСО7, например путем их выведения через почки с мочой. Величина pH плазмы крови зависит от соотношения концентрации растворенного в ней СО2 и ионов бикарбоната. В виде бикарбоната плазмой крови, т. е. в химически связанном состоянии, переносится основное количество углекислого газа —порядка 45 мл/100 мл крови, или до 90 %. Эритроцитами в виде карбаминового соединения с белками гемоглобина транспортируется примерно 2,5 мл/100 мл крови СО2, или 5 %. Транспорт углекислого газа кровью от тканей к легким в указанных формах не связан с явлением насыщения, как при транспорте кислорода, т. е. чем больше образуется углекислого газа, тем большее его количество транспортируется от тканей к легким. Однако между парциальным давлением СО2 в крови и количеством переносимого кровью углекислого газа имеется криволинейная зависимость: кривая диссоциации углекислого газа. 10.4.2.1. Роль эритроцитов в транспорте СО2
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 170; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.143.181 (0.073 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||